

Differential Effector Engagement by Oncogenic KRAS
- PMID: 29444439
- PMCID: PMC6343826
- DOI: 10.1016/j.celrep.2018.01.051
Differential Effector Engagement by Oncogenic KRAS
Abstract
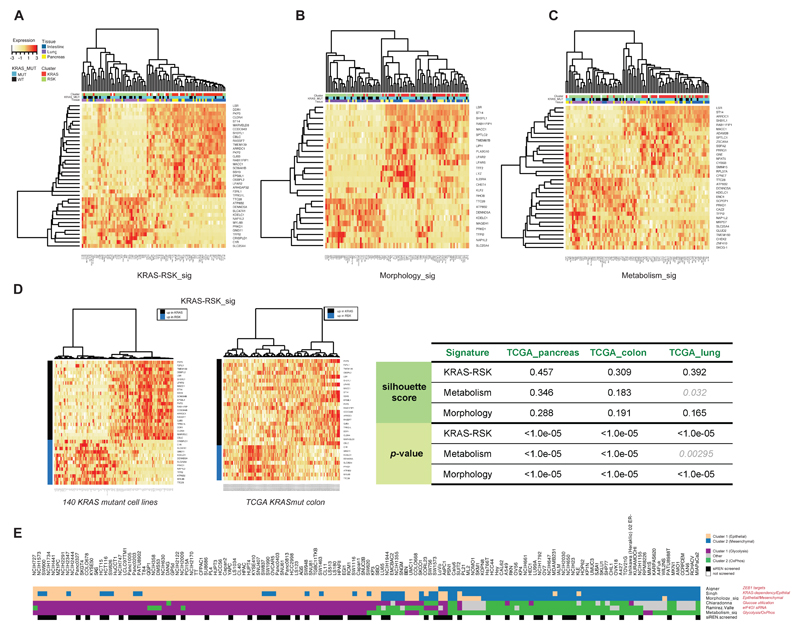
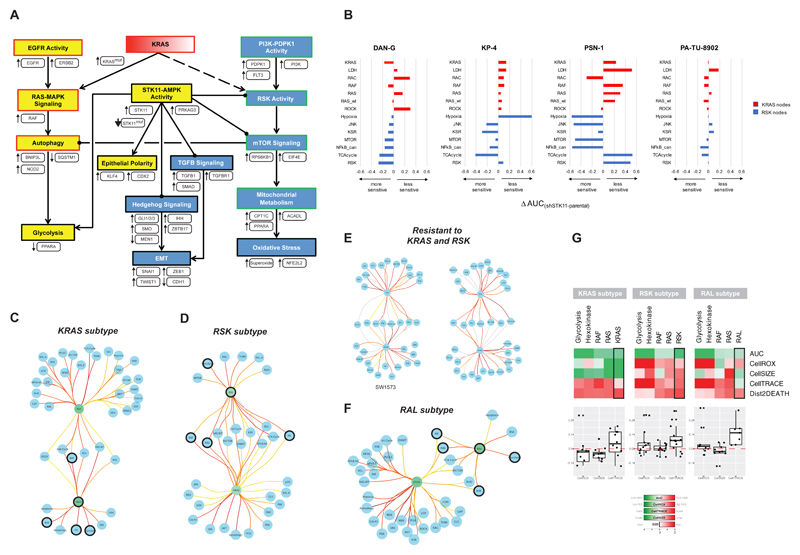
KRAS can bind numerous effector proteins, which activate different downstream signaling events. The best known are RAF, phosphatidylinositide (PI)-3' kinase, and RalGDS families, but many additional direct and indirect effectors have been reported. We have assessed how these effectors contribute to several major phenotypes in a quantitative way, using an arrayed combinatorial siRNA screen in which we knocked down 41 KRAS effectors nodes in 92 cell lines. We show that every cell line has a unique combination of effector dependencies, but in spite of this heterogeneity, we were able to identify two major subtypes of KRAS mutant cancers of the lung, pancreas, and large intestine, which reflect different KRAS effector engagement and opportunities for therapeutic intervention.
Keywords: KRAS; RNAi screen; RSK; paralogs; redundancy.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
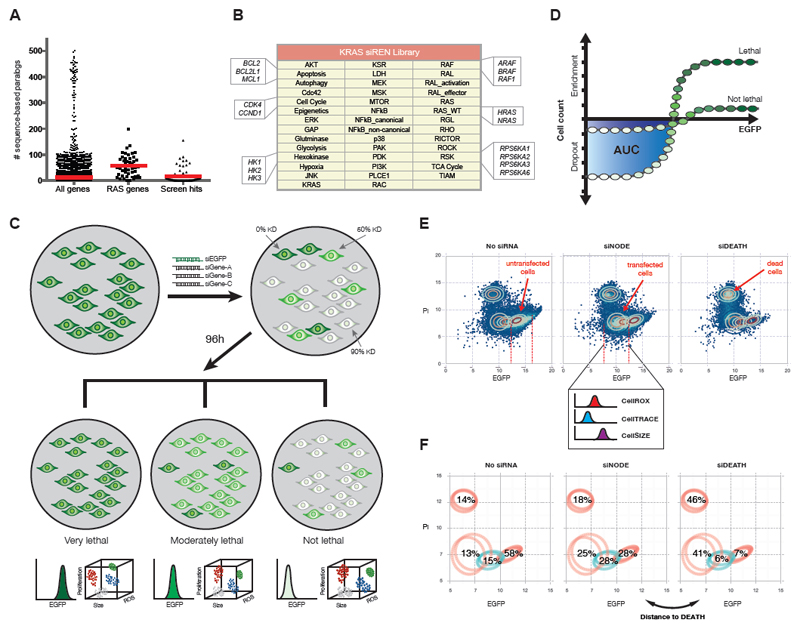
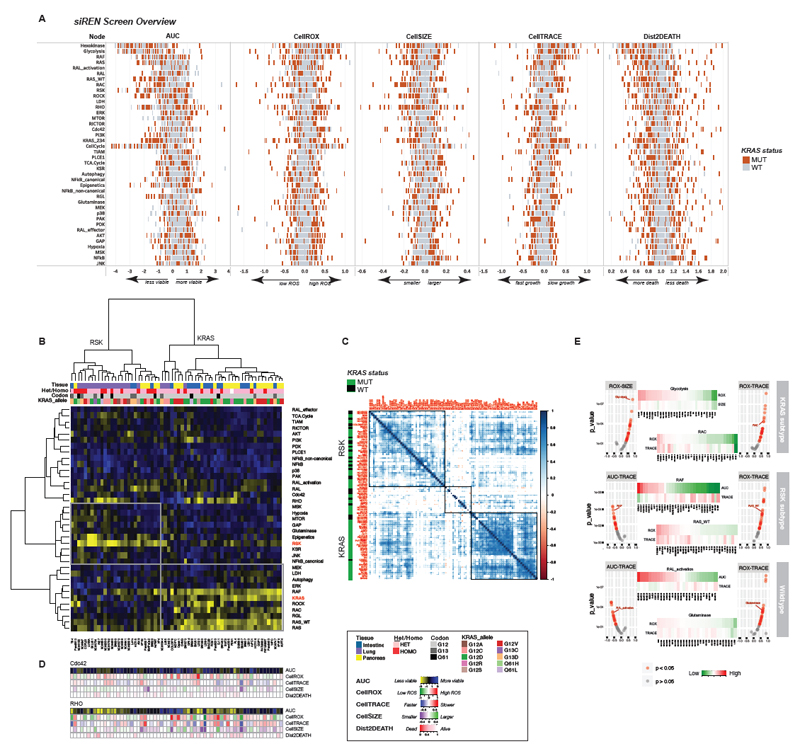
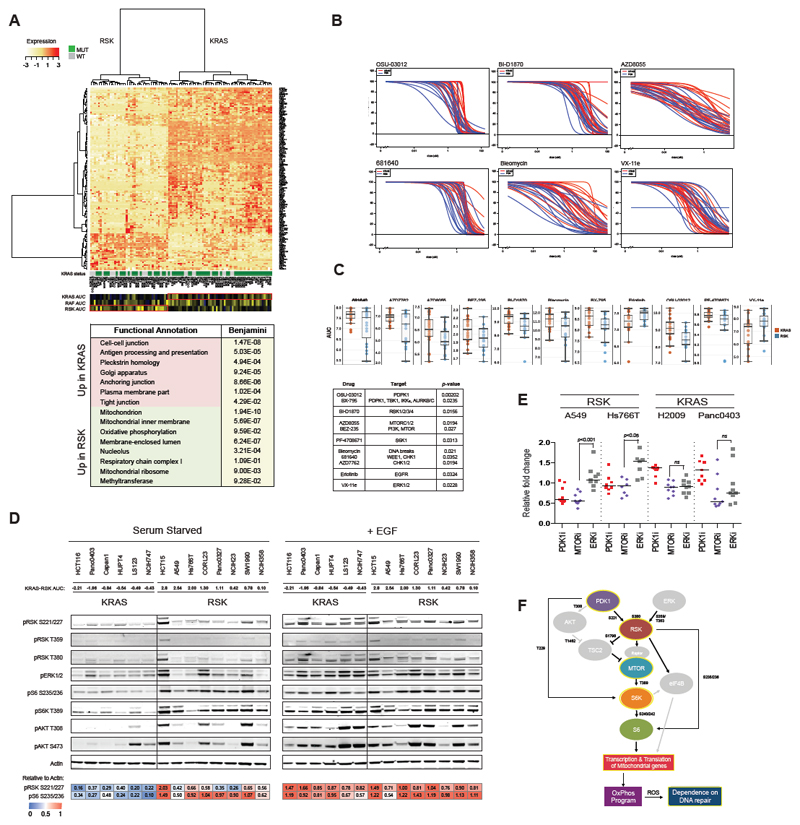
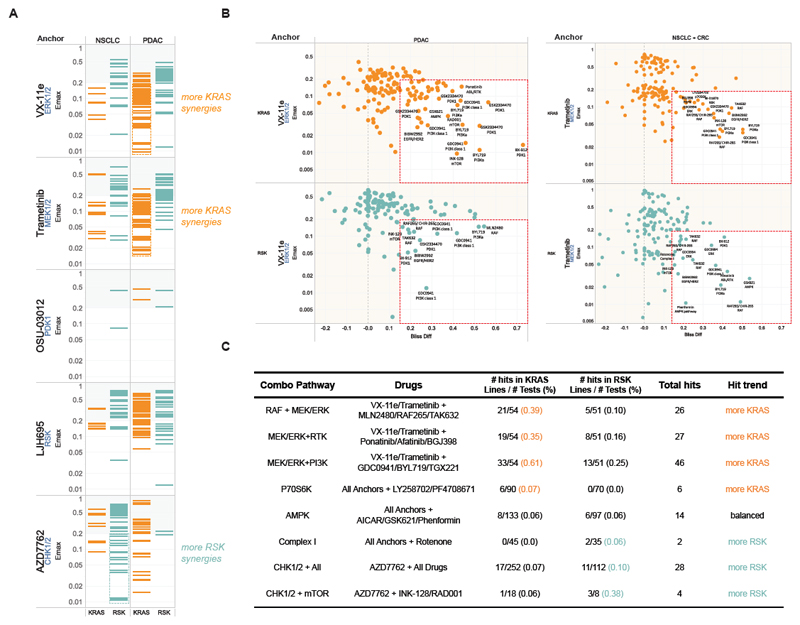
Comment in
-
Mapping Effector-Phenotype Landscapes in KRAS-Driven Cancers.Trends Cancer. 2018 May;4(5):333-335. doi: 10.1016/j.trecan.2018.02.004. Epub 2018 Mar 7. Trends Cancer. 2018. PMID: 29709255
References
-
- Chiaradonna F, Sacco E, Manzoni R, Giorgio M, Vanoni M, Alberghina L. Ras-dependent carbon metabolism and transformation in mouse fibroblasts. Oncogene. 2006;25:5391–5404. - PubMed
-
- Cunningham JT, Rodgers JT, Arlow DH, Vazquez F, Mootha VK, Puigserver P. mTOR controls mitochondrial oxidative function through a YY1-PGC-1alpha transcriptional complex. Nature. 2007;450:736–740. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
