

Pyruvate kinase M2 regulates photoreceptor structure, function, and viability
- PMID: 29445082
- PMCID: PMC5833680
- DOI: 10.1038/s41419-018-0296-4
Pyruvate kinase M2 regulates photoreceptor structure, function, and viability
Abstract
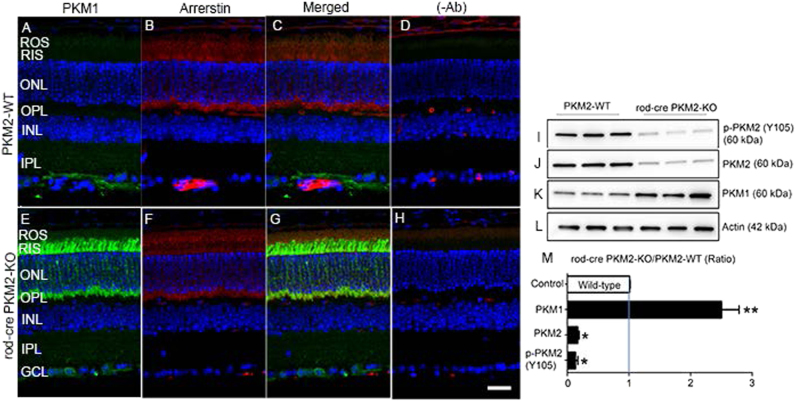
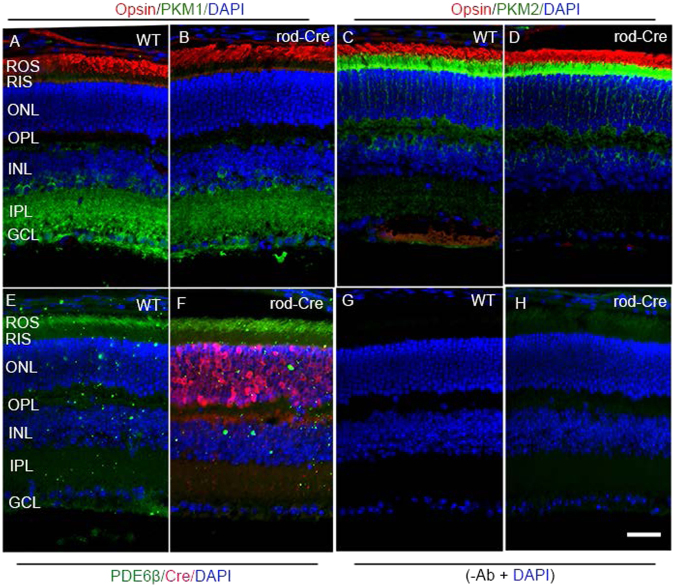
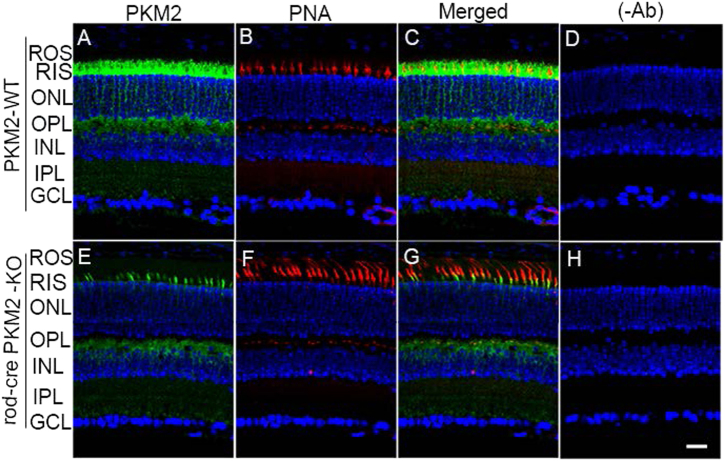
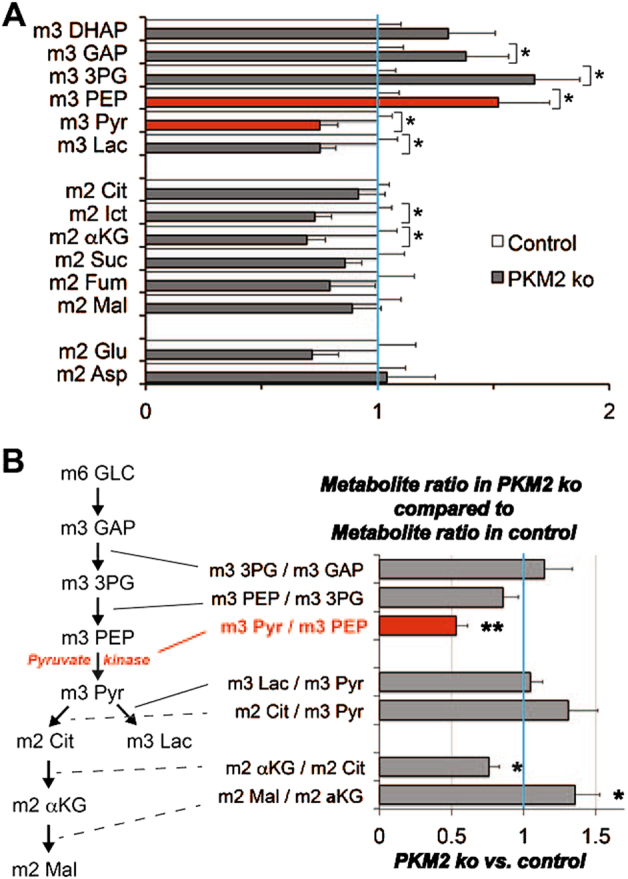
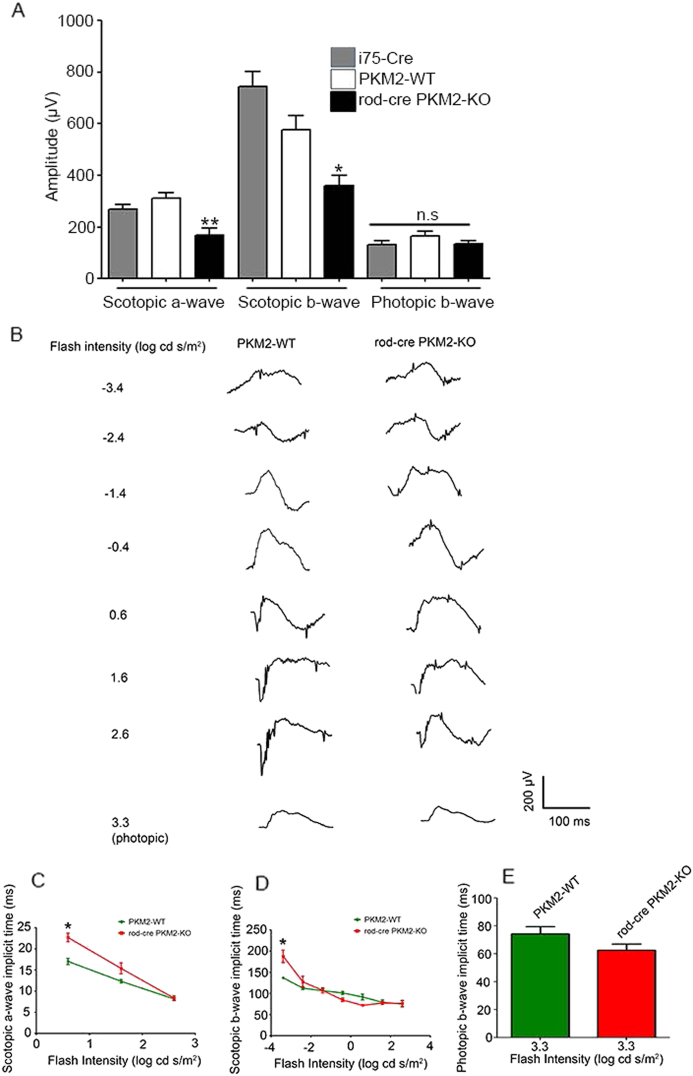
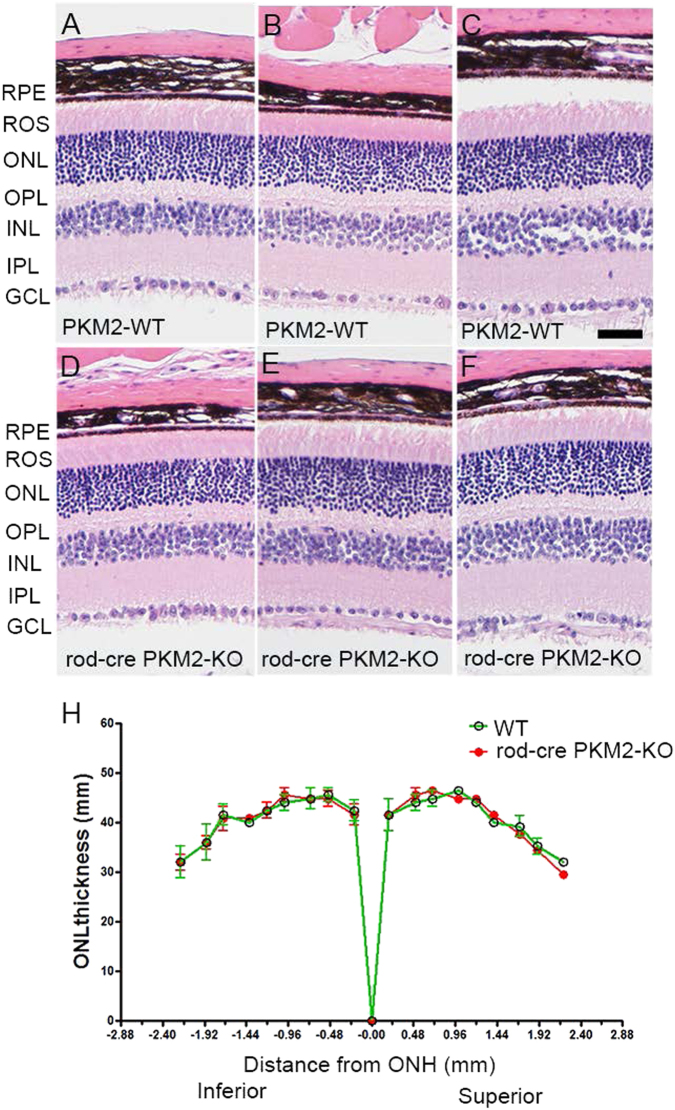
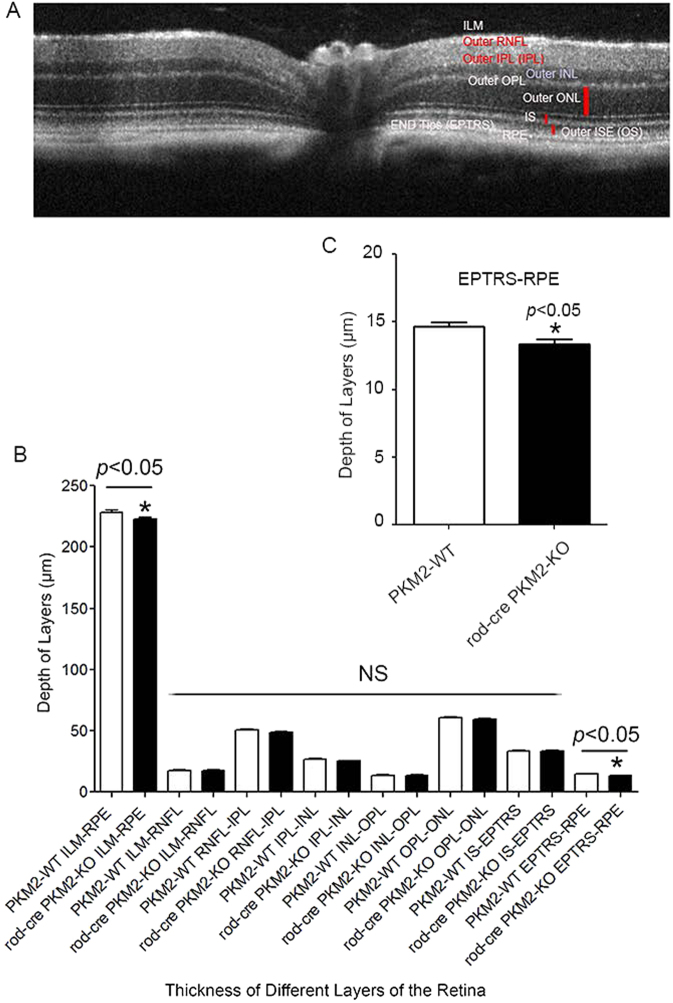
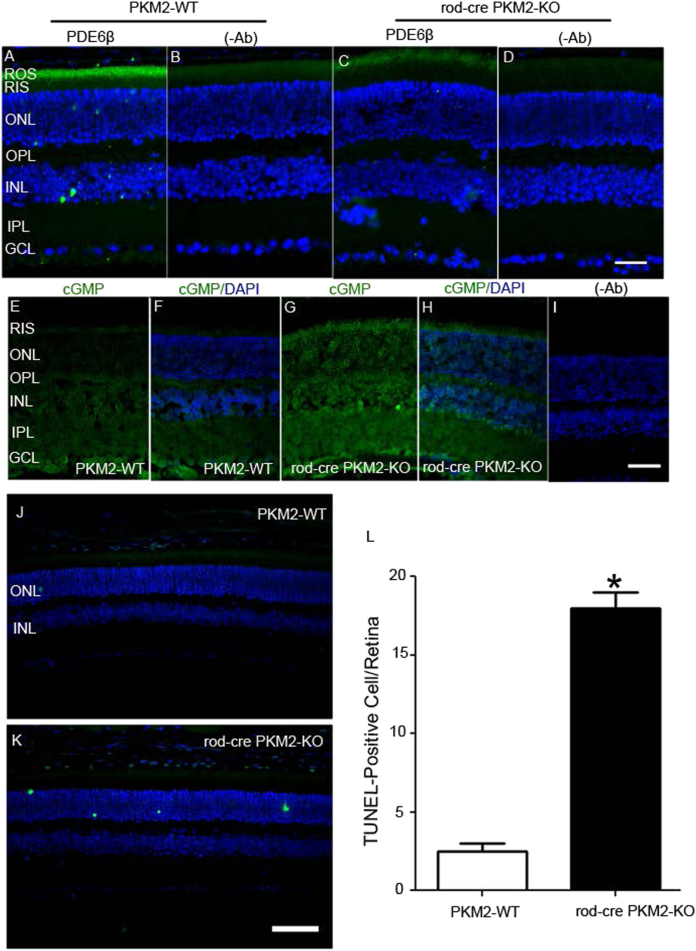
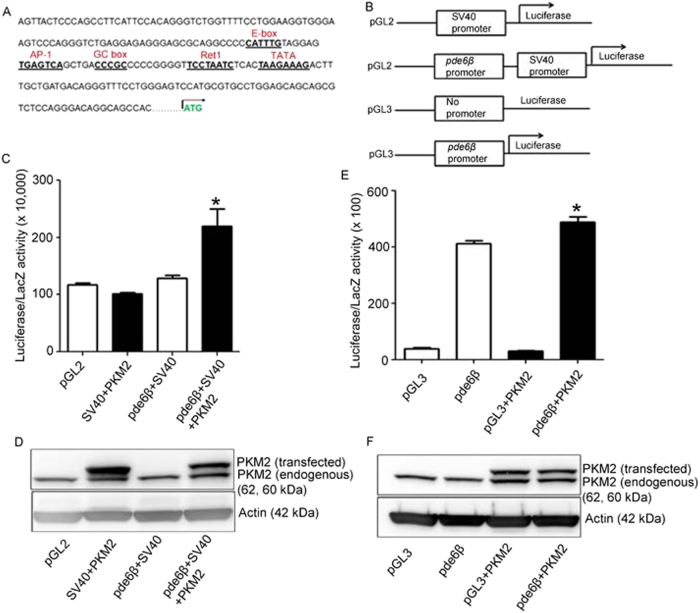
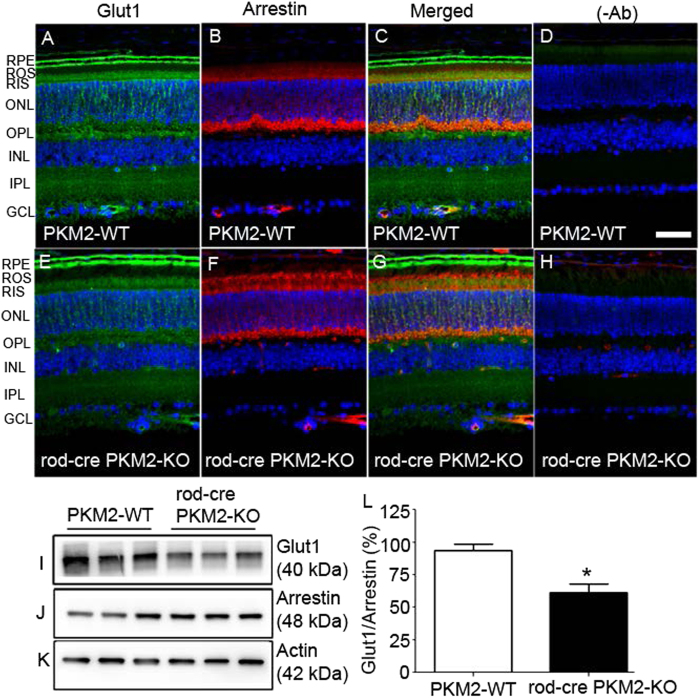
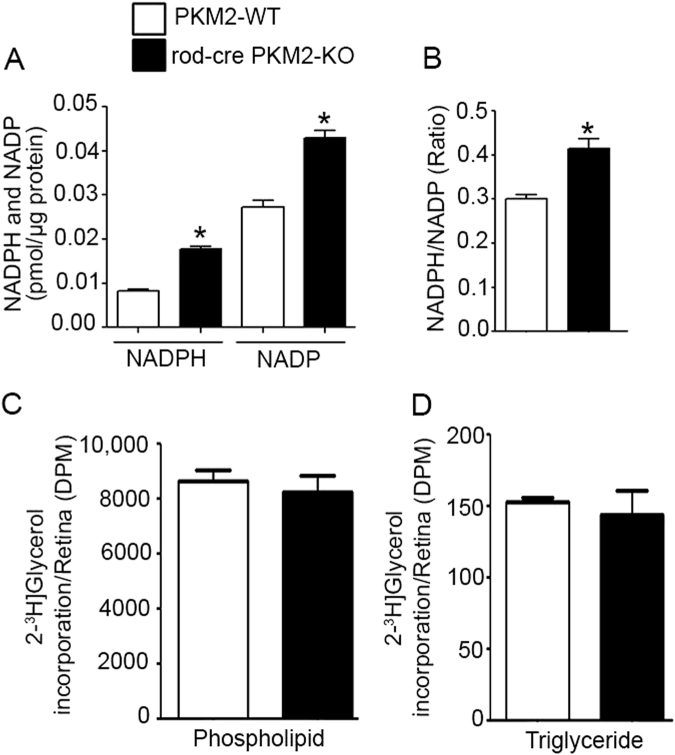
Pyruvate kinase M2 (PKM2) is a glycolytic enzyme that is expressed in cancer cells. Its role in tumor metabolism is not definitively established, but investigators have suggested that regulation of PKM2 activity can cause accumulation of glycolytic intermediates and increase flux through the pentose phosphate pathway. Recent evidence suggests that PKM2 also may have non-metabolic functions, including as a transcriptional co-activator in gene regulation. We reported previously that PKM2 is abundant in photoreceptor cells in mouse retinas. In the present study, we conditionally deleted PKM2 (rod-cre PKM2-KO) in rod photoreceptors and found that the absence of PKM2 causes increased expression of PKM1 in rods. Analysis of metabolic flux from U-13C glucose shows that rod-cre PKM2-KO retinas accumulate glycolytic intermediates, consistent with an overall reduction in the amount of pyruvate kinase activity. Rod-cre PKM2-KO mice also have an increased NADPH availability could favor lipid synthesis, but we found no difference in phospholipid synthesis between rod-cre PKM2 KO and PKM2-positive controls. As rod-cre PKM2-KO mice aged, we observed a significant loss of rod function, reduced thickness of the photoreceptor outer segment layer, and reduced expression of photoreceptor proteins, including PDE6β. The rod-cre PKM2-KO retinas showed greater TUNEL staining than wild-type retinas, indicating a slow retinal degeneration. In vitro analysis showed that PKM2 can regulate transcriptional activity from the PDE6β promoter in vitro. Our findings indicate that both the metabolic and transcriptional regulatory functions of PKM2 may contribute to photoreceptor structure, function, and viability.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Warburg O. On respiratory impairment in cancer cells. Science. 1956;124:269–270. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
