

Integrins promote axonal regeneration after injury of the nervous system
- PMID: 29446228
- PMCID: PMC6055631
- DOI: 10.1111/brv.12398
Integrins promote axonal regeneration after injury of the nervous system
Abstract
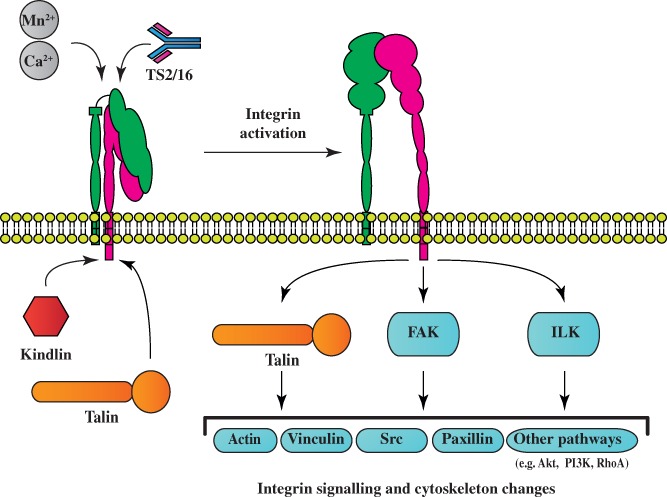
Integrins are cell surface receptors that form the link between extracellular matrix molecules of the cell environment and internal cell signalling and the cytoskeleton. They are involved in several processes, e.g. adhesion and migration during development and repair. This review focuses on the role of integrins in axonal regeneration. Integrins participate in spontaneous axonal regeneration in the peripheral nervous system through binding to various ligands that either inhibit or enhance their activation and signalling. Integrin biology is more complex in the central nervous system. Integrins receptors are transported into growing axons during development, but selective polarised transport of integrins limits the regenerative response in adult neurons. Manipulation of integrins and related molecules to control their activation state and localisation within axons is a promising route towards stimulating effective regeneration in the central nervous system.
Keywords: axon regeneration; integrin; kindlin; receptor activation state; selective polarised transport; traumatic injury of the nervous system.
© 2018 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures



References
-
- Andressen, C. , Adrian, S. , Fässler, R. , Arnhold, S. & Addicks, K. (2005). The contribution of beta1 integrins to neuronal migration and differentiation depends on extracellular matrix molecules. European Journal of Cell Biology 84, 973–982. - PubMed
-
- Andrews, M. R. , Czvitkovich, S. , Dassie, E. , Vogelaar, C. F. , Faissner, A. , Blits, B. , Gage, F. H. , Ffrench‐Constant, C. & Fawcett, J. W. (2009). Alpha9 integrin promotes neurite outgrowth on tenascin‐C and enhances sensory axon regeneration. The Journal of Neuroscience 29, 5546–5557. - PMC - PubMed
-
- Andrews, M. R. , Soleman, S. , Cheah, M. , Tumbarello, D. A. , Mason, M. R. J. , Moloney, E. , Verhaagen, J. , Bensadoun, J.‐C. , Schneider, B. , Aebischer, P. & Fawcett, J. W. (2016). Axonal localization of integrins in the CNS is neuronal type and age dependent. eNeuro 3(4), ENEURO.0029–16.2016. - PMC - PubMed
-
- Arlotta, P. , Molyneaux, B. J. , Chen, J. , Inoue, J. , Kominami, R. & Macklis, J. D. (2005). Neuronal subtype‐specific genes that control corticospinal motor neuron development in vivo. Neuron 45, 207–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
