

Human La binds mRNAs through contacts to the poly(A) tail
- PMID: 29447394
- PMCID: PMC5934636
- DOI: 10.1093/nar/gky090
Human La binds mRNAs through contacts to the poly(A) tail
Abstract
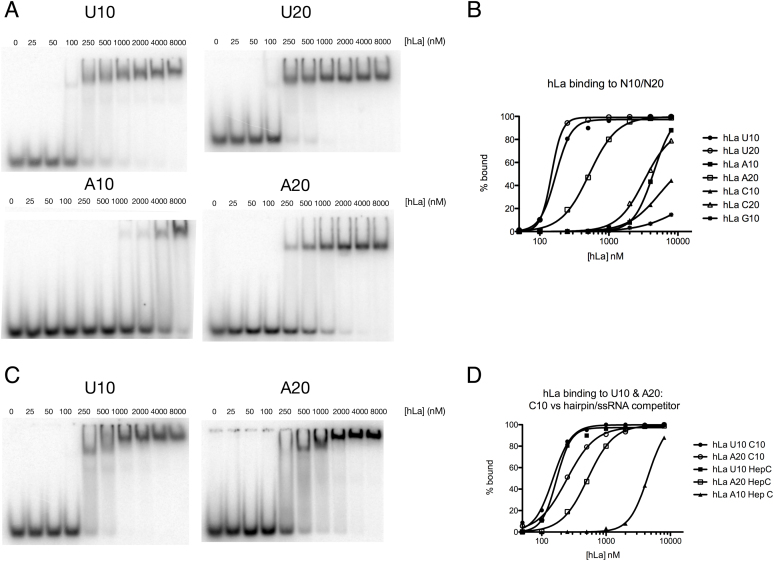
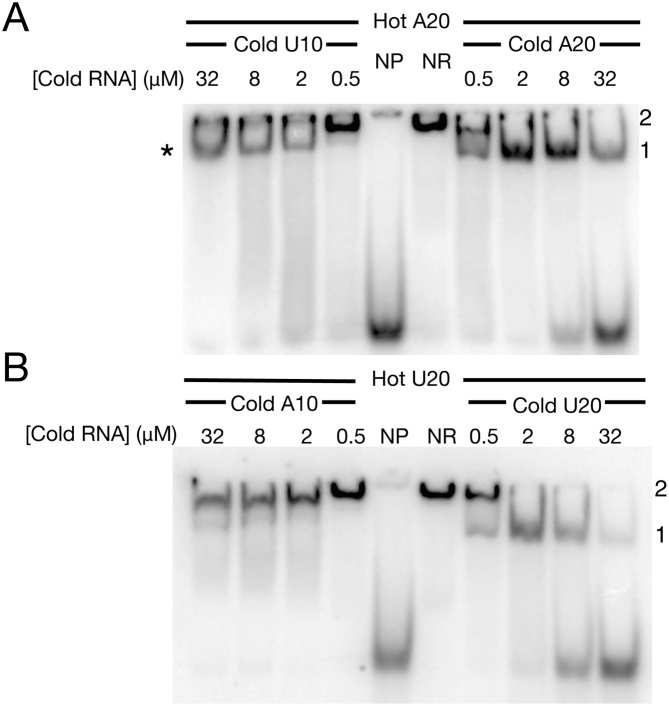
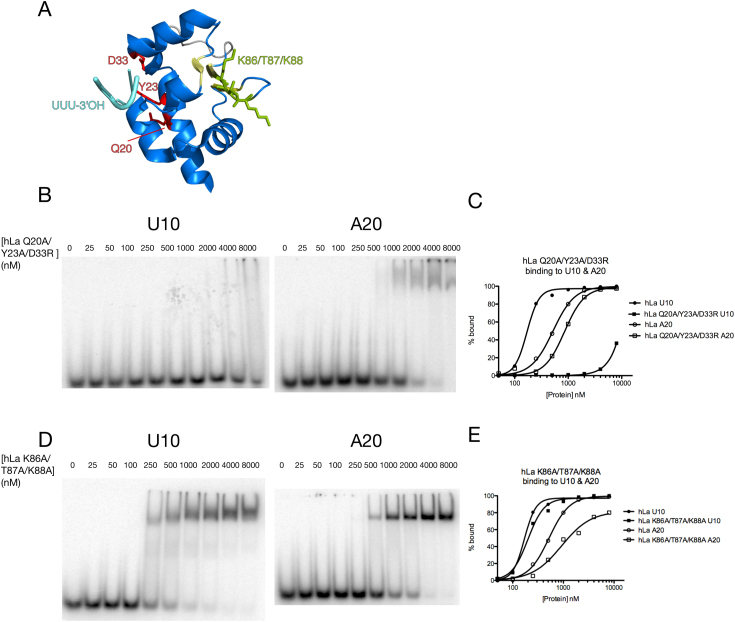
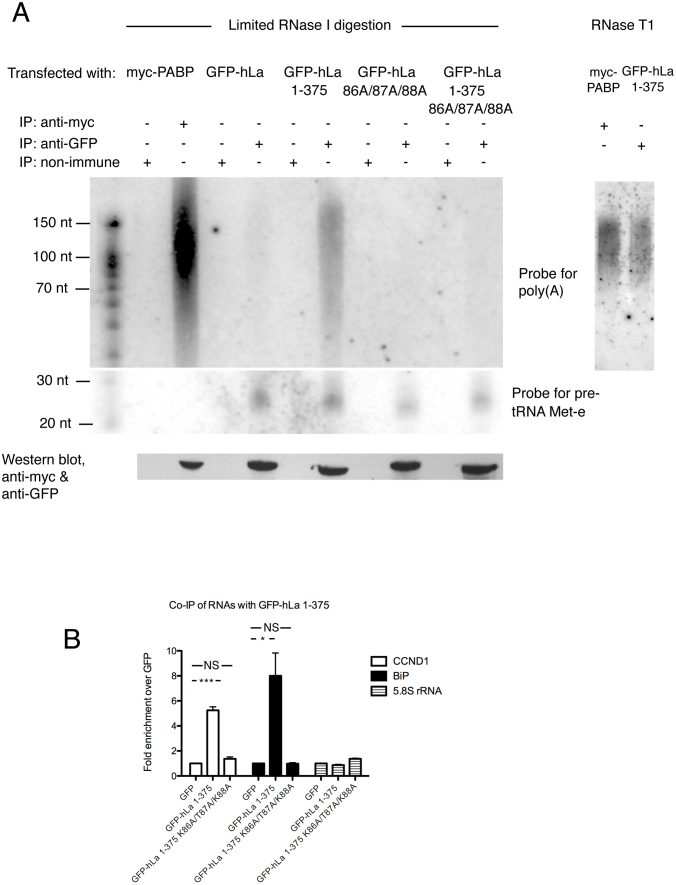
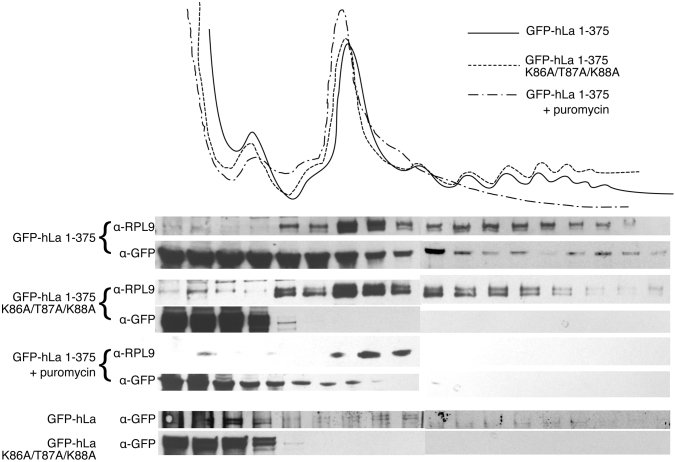
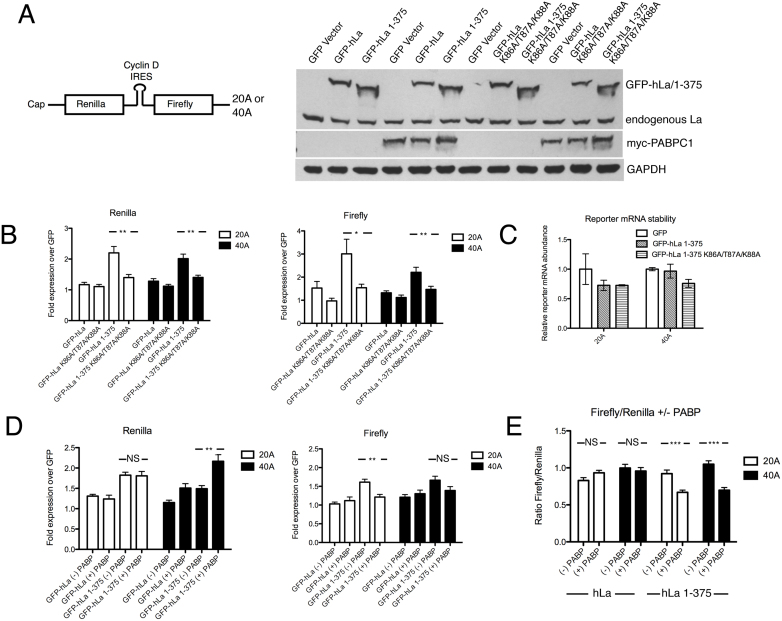
In addition to a role in the processing of nascent RNA polymerase III transcripts, La proteins are also associated with promoting cap-independent translation from the internal ribosome entry sites of numerous cellular and viral coding RNAs. La binding to RNA polymerase III transcripts via their common UUU-3'OH motif is well characterized, but the mechanism of La binding to coding RNAs is poorly understood. Using electromobility shift assays and cross-linking immunoprecipitation, we show that in addition to a sequence specific UUU-3'OH binding mode, human La exhibits a sequence specific and length dependent poly(A) binding mode. We demonstrate that this poly(A) binding mode uses the canonical nucleic acid interaction winged helix face of the eponymous La motif, previously shown to be vacant during uridylate binding. We also show that cytoplasmic, but not nuclear La, engages poly(A) RNA in human cells, that La entry into polysomes utilizes the poly(A) binding mode, and that La promotion of translation from the cyclin D1 internal ribosome entry site occurs in competition with cytoplasmic poly(A) binding protein (PABP). Our data are consistent with human La functioning in translation through contacts to the poly(A) tail.
Figures
References
-
- Mattioli M., Reichlin M.. Heterogeneity of RNA protein antigens reactive with sera of patients with systemic lupus erythematosus. Description of a cytoplasmic nonribosomal antigen. Arthritis Rheum. 1974; 17:421–429. - PubMed
-
- Lerner M.R., Boyle J.A., Hardin J.A., Steitz J.A.. Two novel classes of small ribonucleoproteins detected by antibodies associated with lupus erythematosus. Science. 1981; 211:400–402. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
