

Place learning overrides innate behaviors in Drosophila
- PMID: 29449456
- PMCID: PMC5817280
- DOI: 10.1101/lm.046136.117
Place learning overrides innate behaviors in Drosophila
Abstract
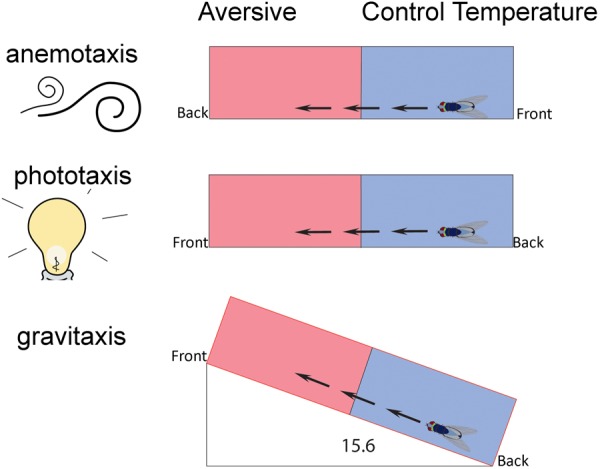
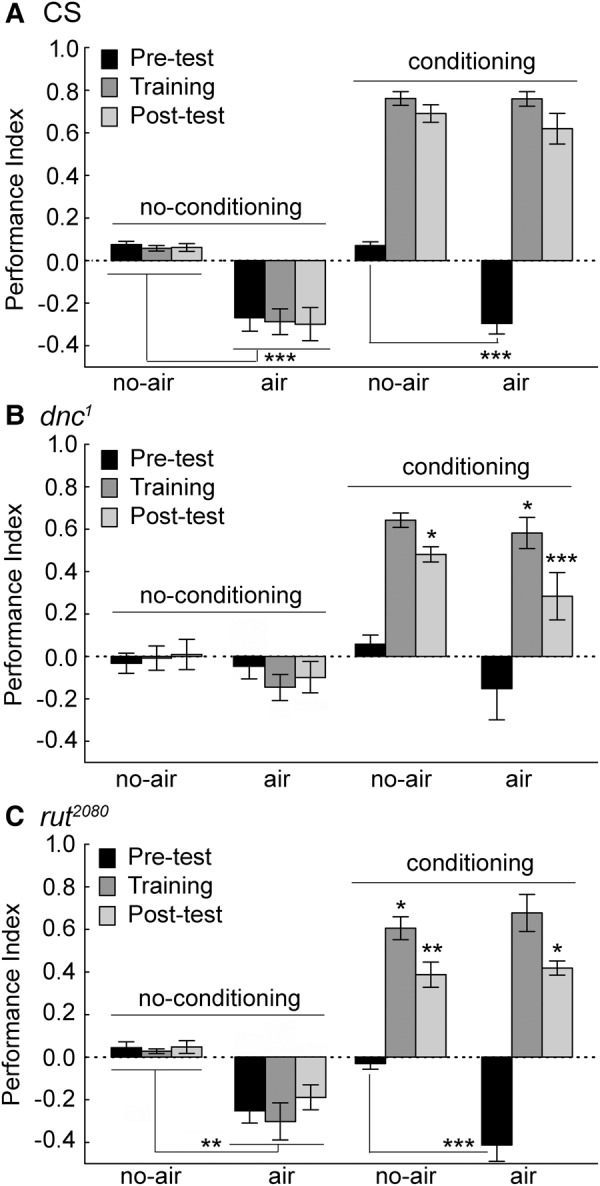
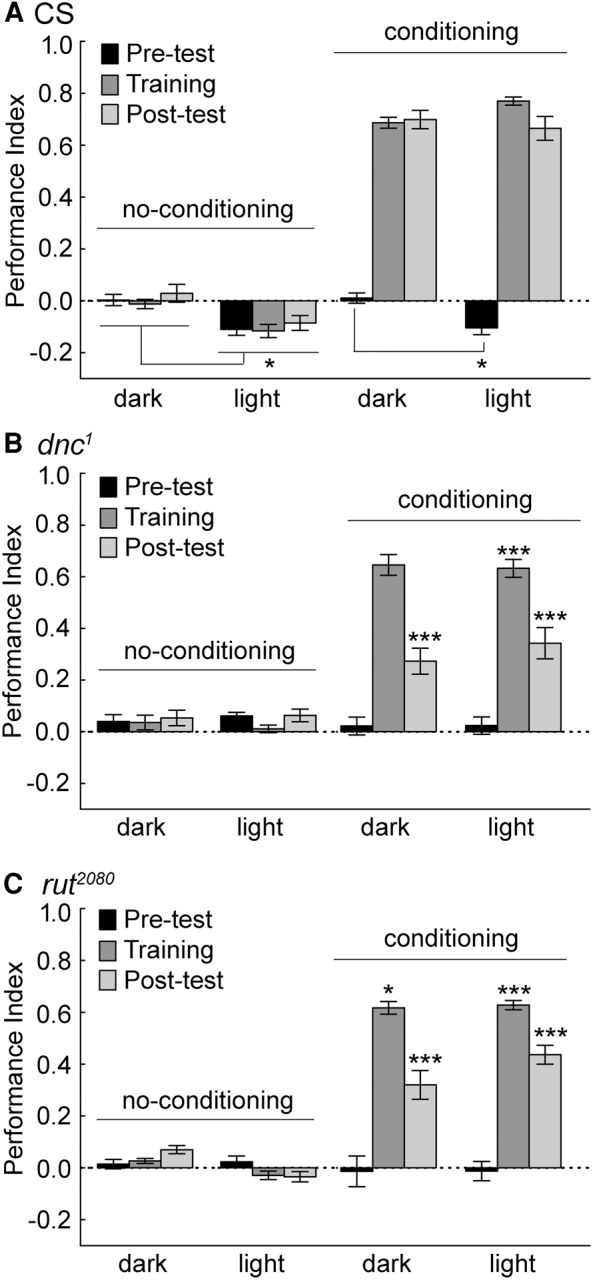
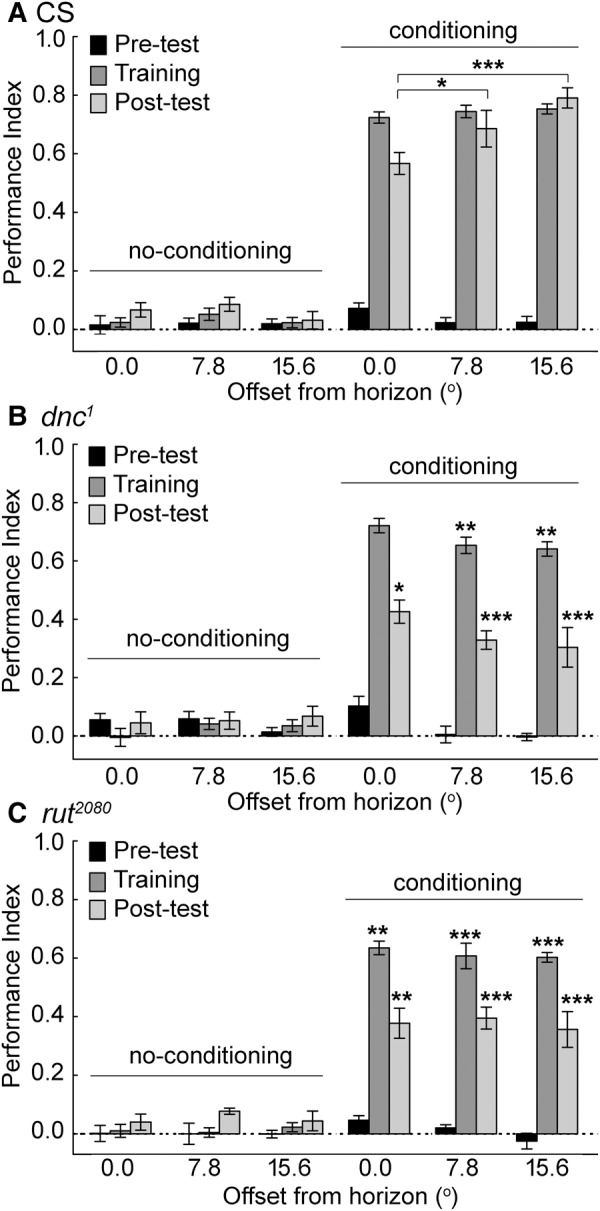
Animals in a natural environment confront many sensory cues. Some of these cues bias behavioral decisions independent of experience, and action selection can reveal a stimulus-response (S-R) connection. However, in a changing environment it would be a benefit for an animal to update behavioral action selection based on experience, and learning might modify even strong S-R relationships. How animals use learning to modify S-R relationships is a largely open question. Three sensory stimuli, air, light, and gravity sources were presented to individual Drosophila melanogaster in both naïve and place conditioning situations. Flies were tested for a potential modification of the S-R relationships of anemotaxis, phototaxis, and negative gravitaxis by a contingency that associated place with high temperature. With two stimuli, significant S-R relationships were abandoned when the cue was in conflict with the place learning contingency. The role of the dunce (dnc) cAMP-phosphodiesterase and the rutabaga (rut) adenylyl cyclase were examined in all conditions. Both dnc1 and rut2080 mutant flies failed to display significant S-R relationships with two attractive cues, and have characteristically lower conditioning scores under most conditions. Thus, learning can have profound effects on separate native S-R relationships in multiple contexts, and mutation of the dnc and rut genes reveal complex effects on behavior.
© 2018 Baggett et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Armstrong JD, Texada MJ, Munjaal R, Baker DA, Beckingham KM. 2006. Gravitaxis in Drosophila melanogaster: a forward genetic screen. Genes Brain Behav 5: 222–239. - PubMed
-
- Budick SA, Reiser MB, Dickinson MH. 2007. The role of visual and mechanosensory cues in structuring forward flight in Drosophila melanogaster. J Exp Biol 210: 4092–4103. - PubMed
-
- Chow DM, Frye MA. 2008. Context-dependent olfactory enhancement of optomotor flight control in Drosophila. J Exp Biol 211: 2478–2485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials