

Macrogenomic engineering via modulation of the scaling of chromatin packing density
- PMID: 29450107
- PMCID: PMC5809134
- DOI: 10.1038/s41551-017-0153-2
Macrogenomic engineering via modulation of the scaling of chromatin packing density
Abstract
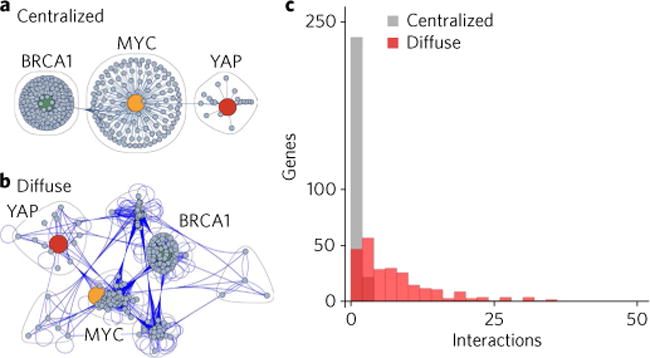
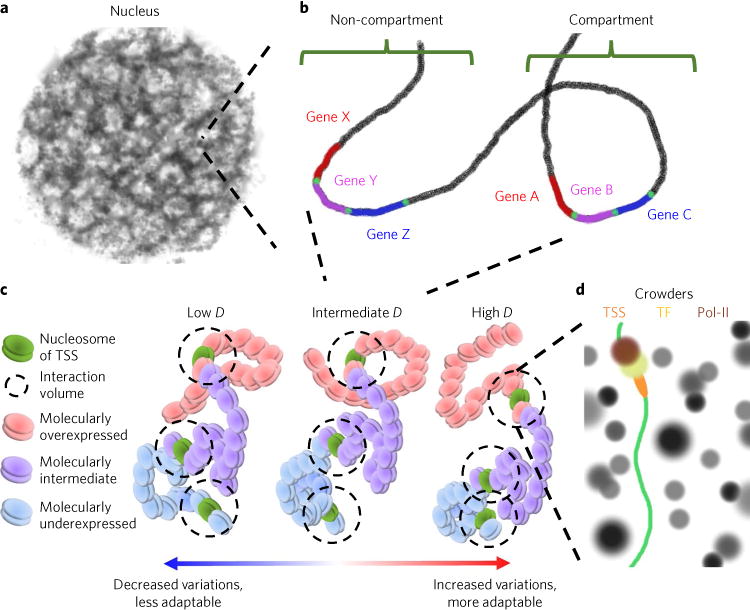
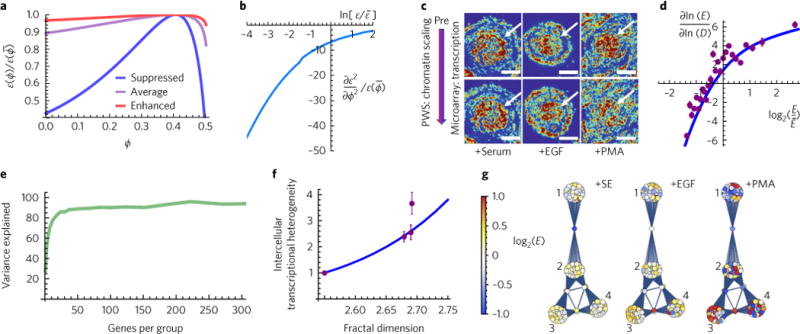
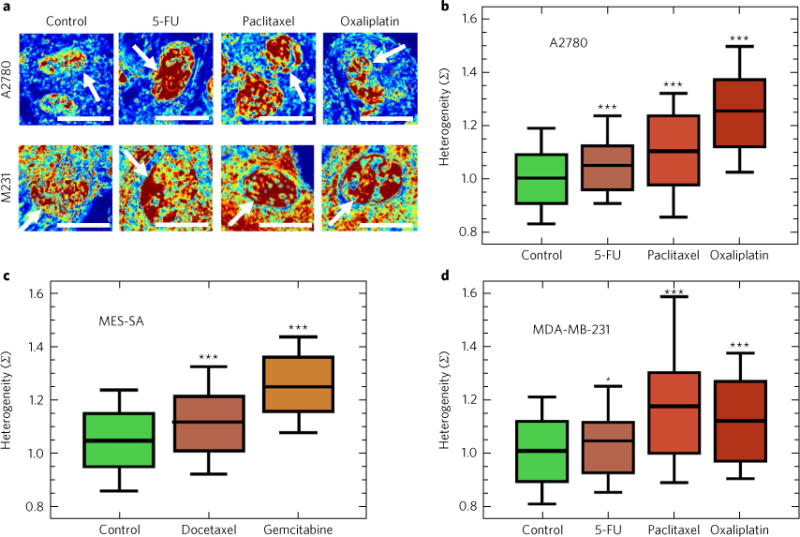
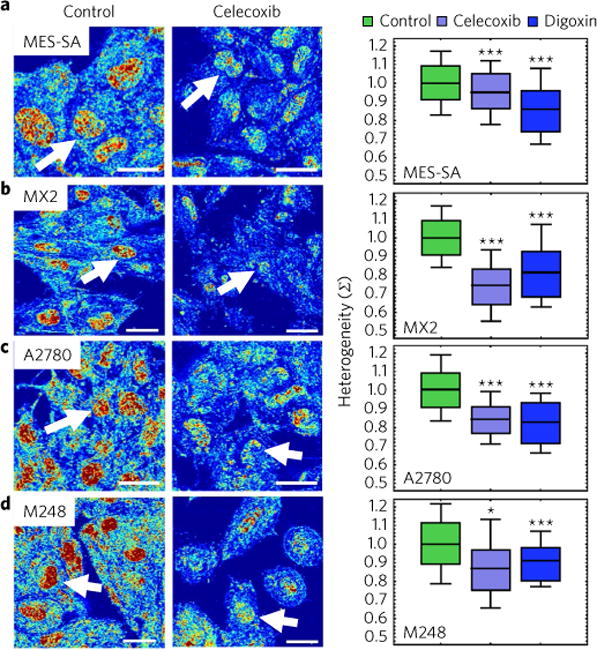
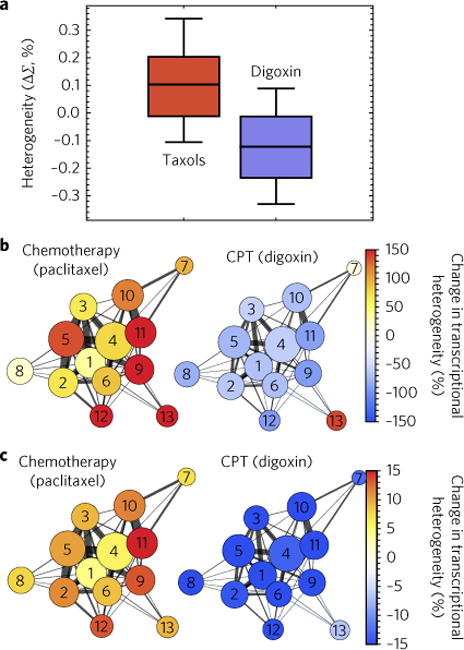
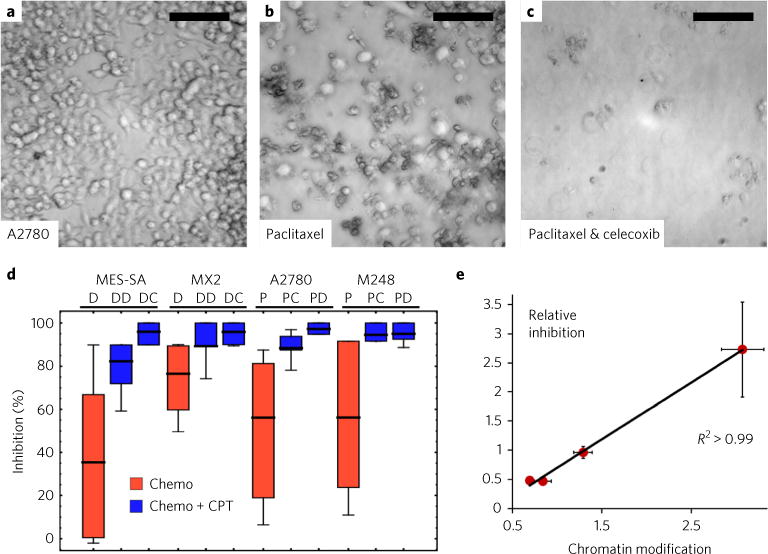
Many human diseases result from the dysregulation of the complex interactions between tens to thousands of genes. However, approaches for the transcriptional modulation of many genes simultaneously in a predictive manner are lacking. Here, through the combination of simulations, systems modelling and in vitro experiments, we provide a physical regulatory framework based on chromatin packing-density heterogeneity for modulating the genomic information space. Because transcriptional interactions are essentially chemical reactions, they depend largely on the local physical nanoenvironment. We show that the regulation of the chromatin nanoenvironment allows for the predictable modulation of global patterns in gene expression. In particular, we show that the rational modulation of chromatin density fluctuations can lead to a decrease in global transcriptional activity and intercellular transcriptional heterogeneity in cancer cells during chemotherapeutic responses to achieve near-complete cancer cell killing in vitro. Our findings represent a 'macrogenomic engineering' approach to modulating the physical structure of chromatin for whole-scale transcriptional modulation.
Conflict of interest statement
Competing interests The authors declare no competing financial interests.
Figures
References
-
- Collins FS. Shattuck lecture—medical and societal consequences of the Human Genome Project. N Engl J Med. 1999;341:28–37. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
