

Glutamatergic Ventral Pallidal Neurons Modulate Activity of the Habenula-Tegmental Circuitry and Constrain Reward Seeking
- PMID: 29452828
- PMCID: PMC5972062
- DOI: 10.1016/j.biopsych.2018.01.003
Glutamatergic Ventral Pallidal Neurons Modulate Activity of the Habenula-Tegmental Circuitry and Constrain Reward Seeking
Abstract
Background: The ability to appropriately integrate and respond to rewarding and aversive stimuli is essential for survival. The ventral pallidum (VP) plays a critical role in processing both rewarding and aversive stimuli. However, the VP is a heterogeneous structure, and how VP subpopulations integrate into larger reward networks to ultimately modulate these behaviors is not known. We identify a noncanonical population of glutamatergic VP neurons that play a unique role in responding to aversive stimuli and constraining inappropriate reward seeking.
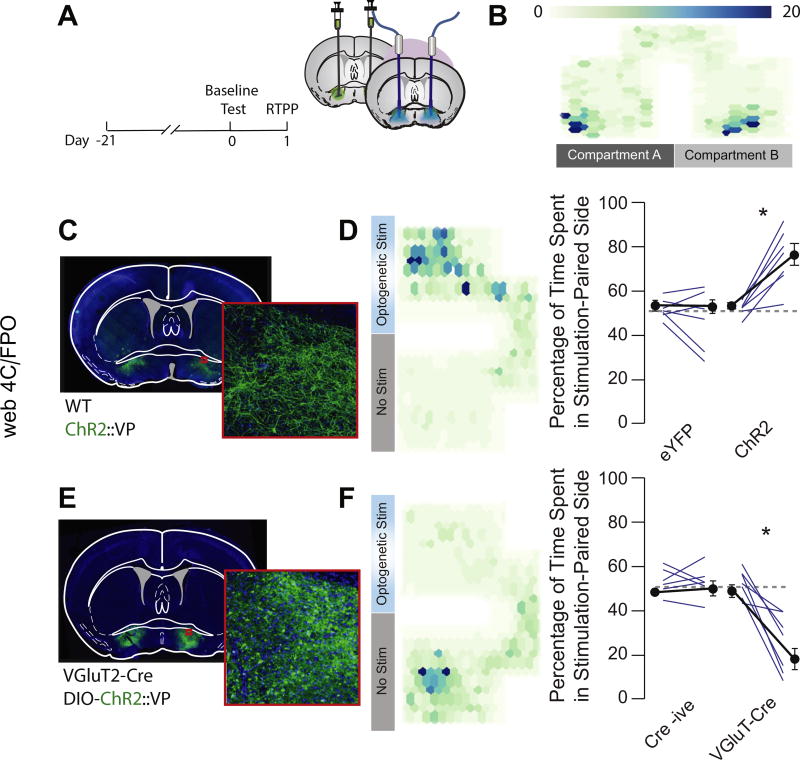
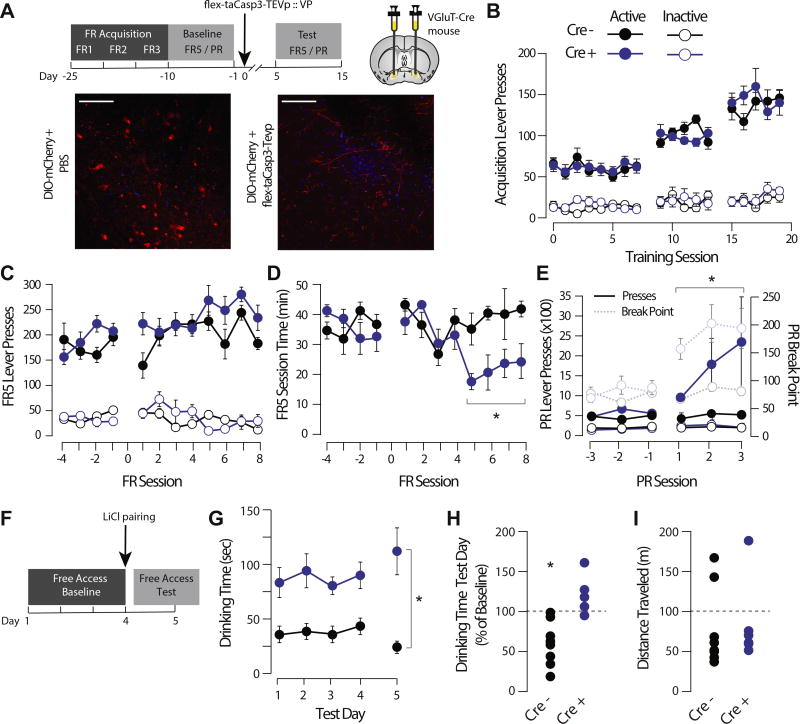
Methods: Using neurochemical, genetic, and electrophysiological approaches, we characterized glutamatergic VP neurons (n = 4-8 mice/group). We performed patch clamp and in vivo electrophysiology recordings in the lateral habenula, rostromedial tegmental nucleus, and ventral tegmental area to determine the effect of glutamatergic VP neuron activation in these target regions (n = 6-10 mice/group). Finally, we selectively optogenetically stimulated glutamatergic VP neurons in a real-time place preference task and ablated these neurons using a virally expressed caspase to determine their necessity for reward seeking.
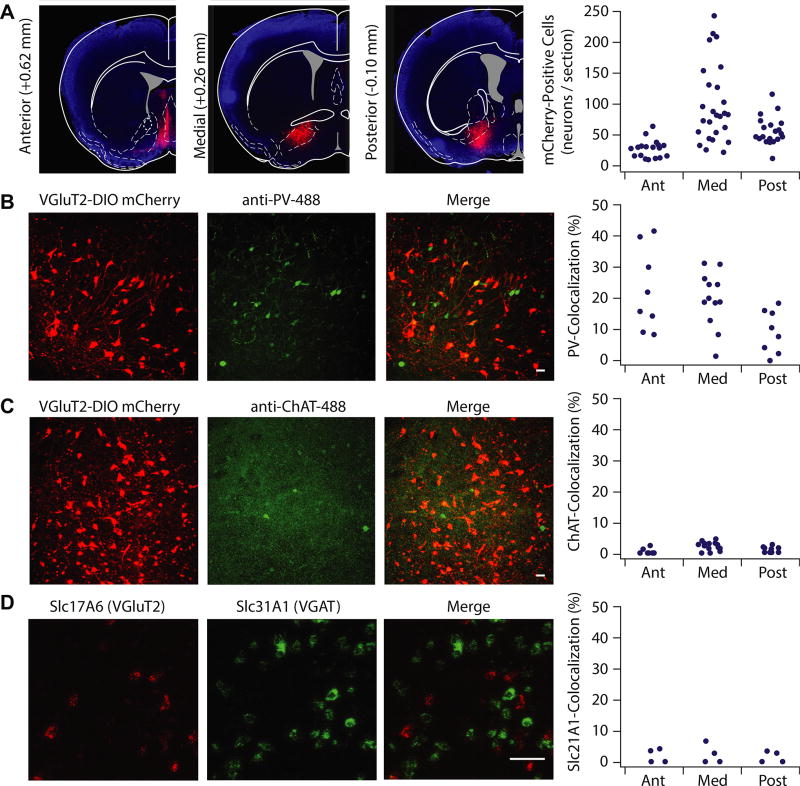
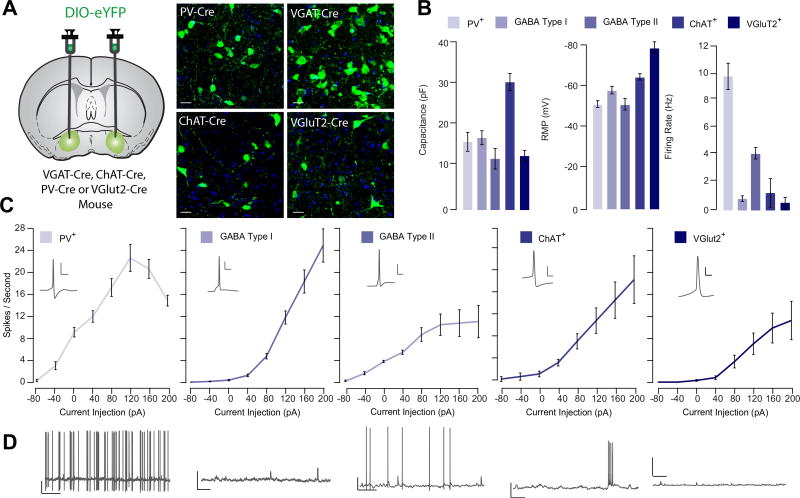
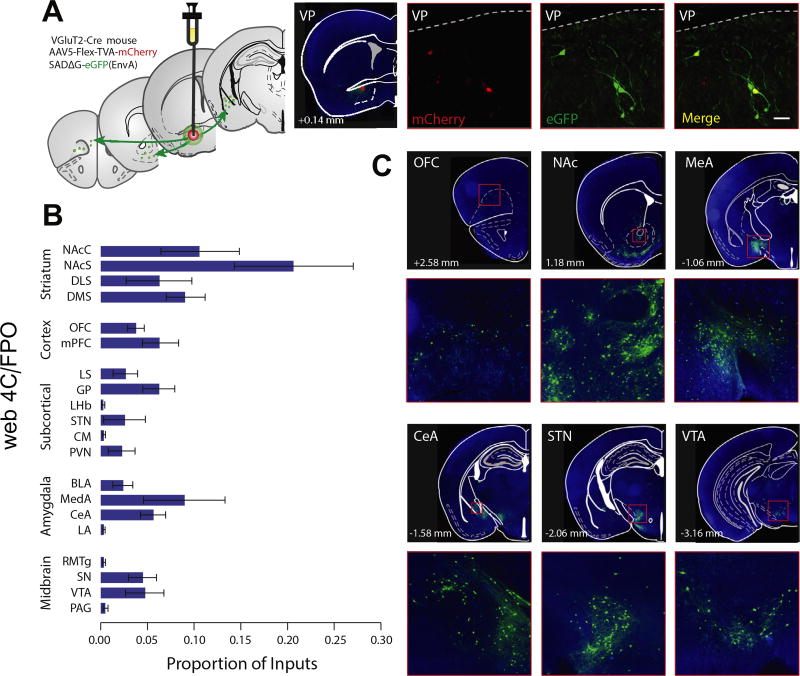
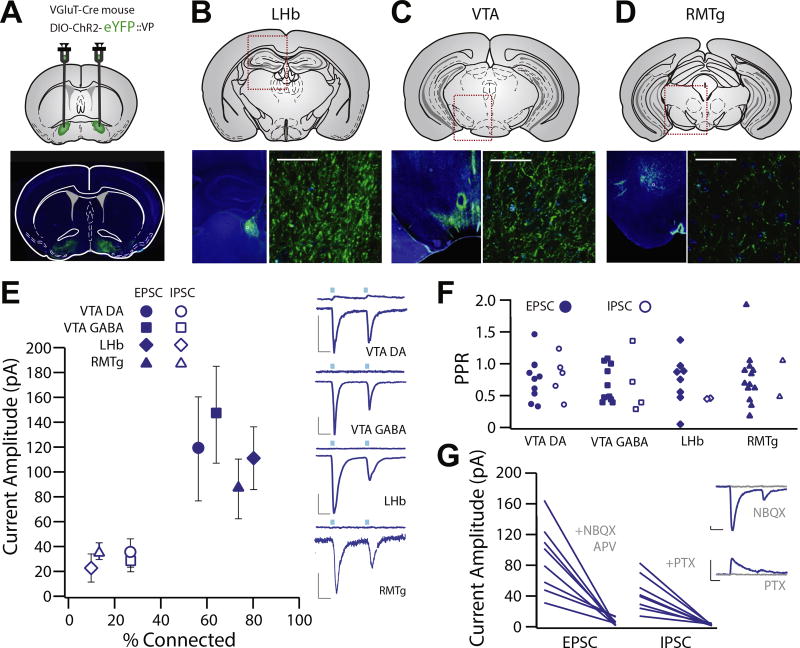
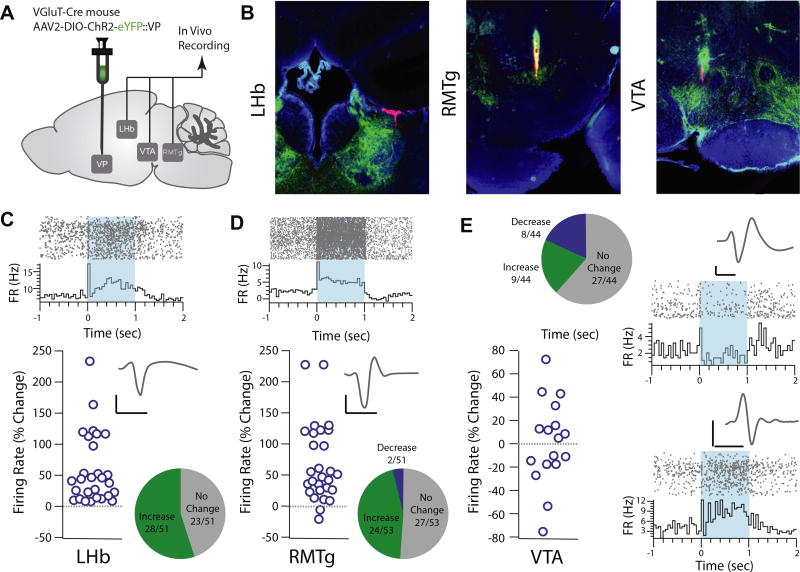
Results: Glutamatergic VP neurons exhibit little overlap with cholinergic or gamma-aminobutyric acidergic markers, the canonical VP subtypes, and exhibit distinct membrane properties. Glutamatergic VP neurons innervate and increase firing activity of the lateral habenula, rostromedial tegmental nucleus, and gamma-aminobutyric acidergic ventral tegmental area neurons. While nonselective optogenetic stimulation of the VP induced a robust place preference, selective activation of glutamatergic VP neurons induced a place avoidance. Viral ablation of glutamatergic VP neurons increased reward responding and abolished taste aversion to sucrose.
Conclusions: Glutamatergic VP neurons constitute a noncanonical subpopulation of VP neurons. These glutamatergic VP neurons increase activity of the lateral habenula, rostromedial tegmental nucleus, and gamma-aminobutyric acidergic ventral tegmental area neurons and adaptively constrain reward seeking.
Keywords: Aversion; Dopamine (DA); Electrophysiology; Gamma-aminobutyric acid (GABA); Rostromedial tegmental nucleus (RMTg); Ventral tegmental area (VTA).
Copyright © 2018 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors report no biomedical financial interests or potential conflicts of interest.
Figures
Comment in
-
The Control of Reward Seeking.Biol Psychiatry. 2018 Jun 15;83(12):981-983. doi: 10.1016/j.biopsych.2018.04.003. Biol Psychiatry. 2018. PMID: 29804588 No abstract available.
References
-
- Haber SN, Groenewegen HJ, Grove EA, Nauta WJ. Efferent connections of the ventral pallidum: Evidence of a dual striato pallidofugal pathway. J Comp Neurol. 1985;235:322–335. - PubMed
-
- Feil J, Sheppard D, Fitzgerald PB, Yucel M, Lubman DI, Bradshaw JL. Addiction, compulsive drug seeking, and the role of frontostriatal mechanisms in regulating inhibitory control. Neurosci Biobehav Rev. 2010;35:248–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
