

A role for tau in learning, memory and synaptic plasticity
- PMID: 29453339
- PMCID: PMC5816660
- DOI: 10.1038/s41598-018-21596-3
A role for tau in learning, memory and synaptic plasticity
Abstract
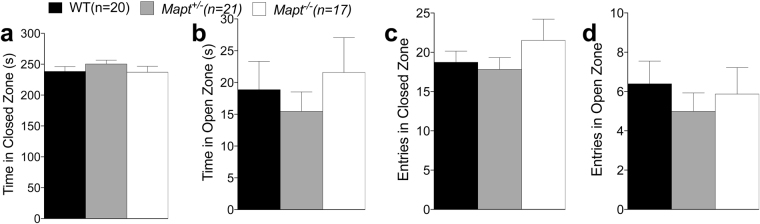
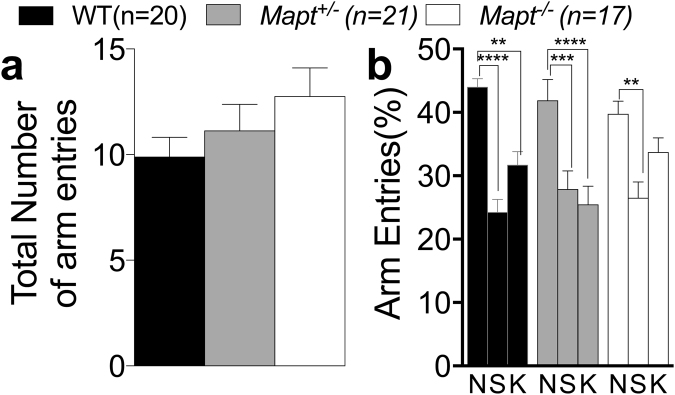
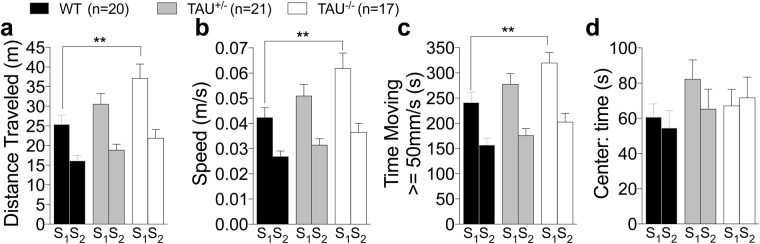
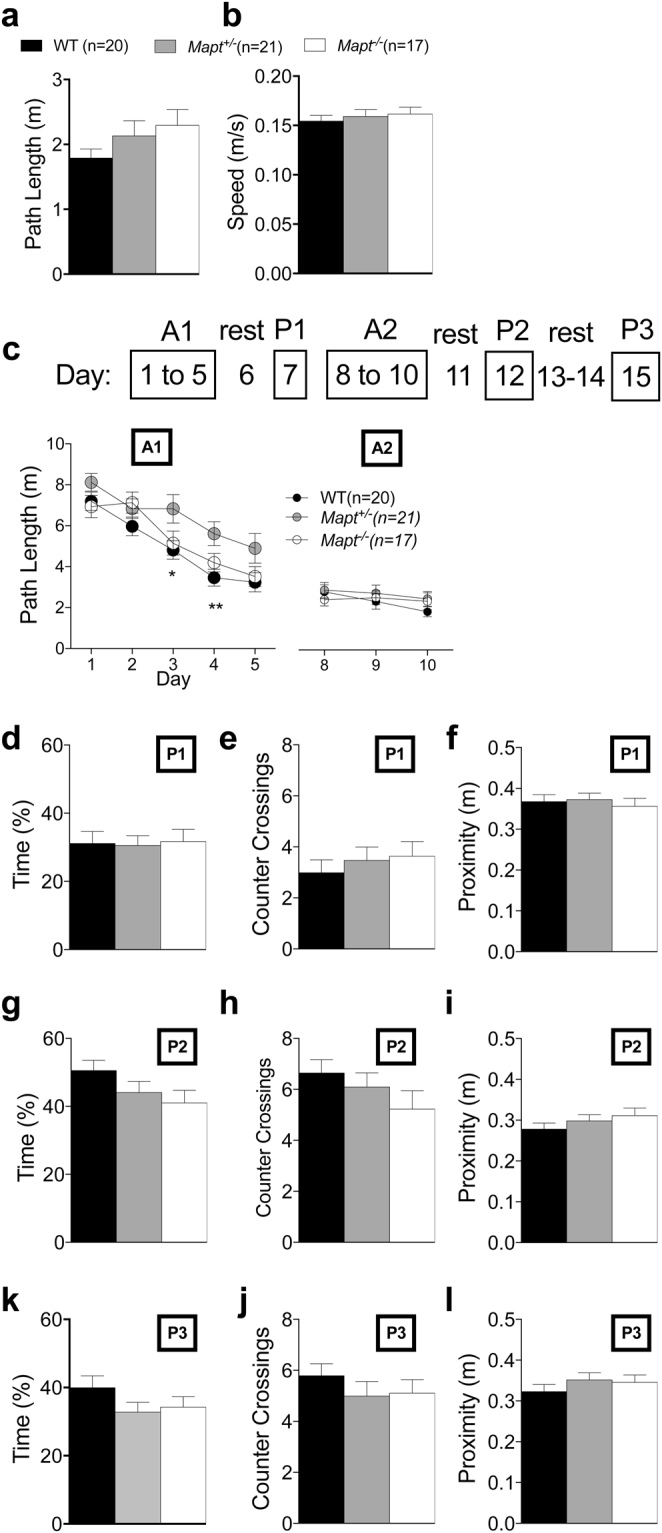
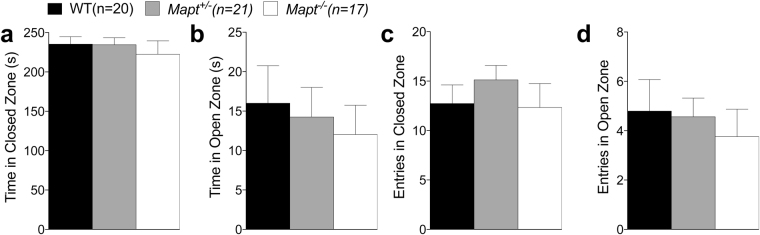
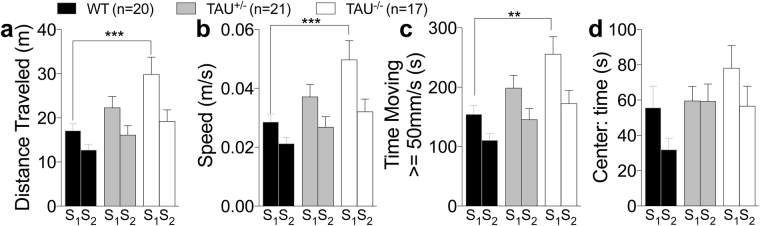
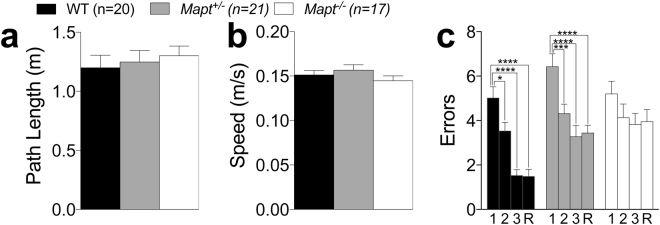
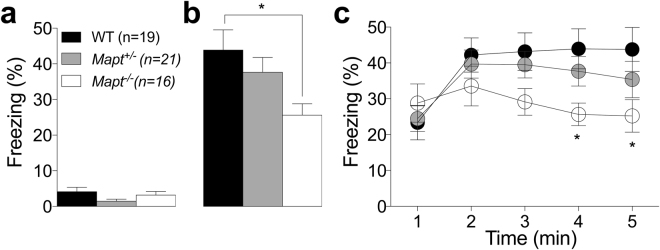
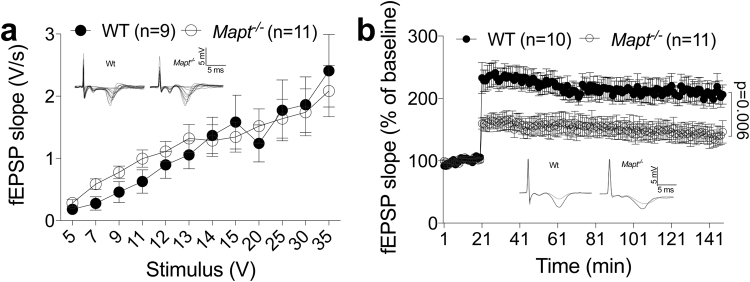
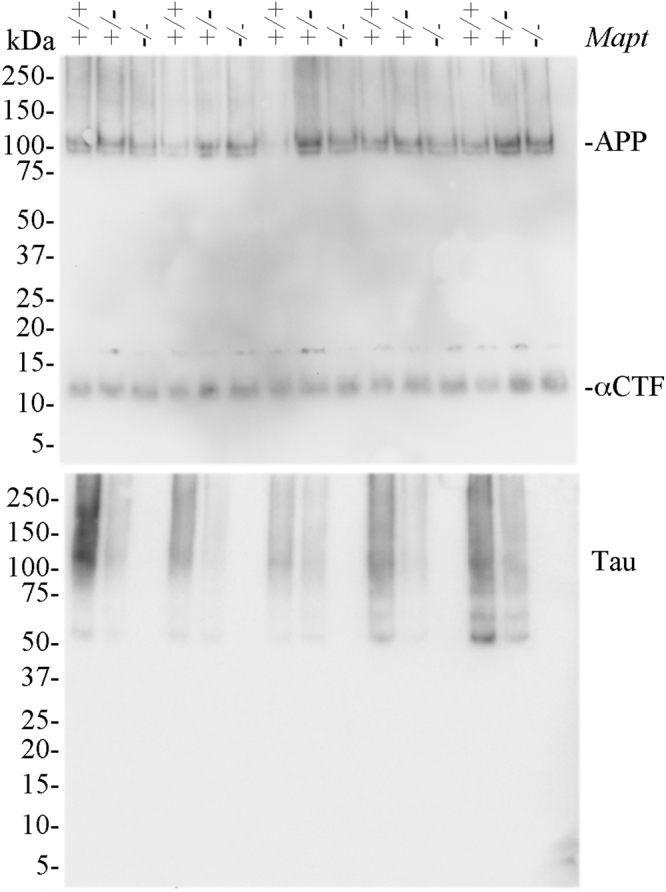
Tau plays a pivotal role in the pathogenesis of neurodegenerative disorders: mutations in the gene encoding for tau (MAPT) are linked to Fronto-temporal Dementia (FTD) and hyper-phosphorylated aggregates of tau forming neurofibrillary tangles (NFTs) that constitute a pathological hallmark of Alzheimer disease (AD) and FTD. Accordingly, tau is a favored therapeutic target for the treatment of these diseases. Given the criticality of tau to dementia's pathogenesis and therapy, it is important to understand the physiological function of tau in the central nervous system. Analysis of Mapt knock out (Mapt-/-) mice has yielded inconsistent results. Some studies have shown that tau deletion does not alter memory while others have described synaptic plasticity and memory alterations in Mapt-/- mice. To help clarifying these contrasting results, we analyzed a distinct Mapt-/- model on a B6129PF3/J genetic background. We found that tau deletion leads to aging-dependent short-term memory deficits, hyperactivity and synaptic plasticity defects. In contrast, Mapt+/- mice only showed a mild short memory deficit in the novel object recognition task. Thus, while tau is important for normal neuronal functions underlying learning and memory, partial reduction of tau expression may have fractional deleterious effects.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
