

Mechanism for initiation of food allergy: Dependence on skin barrier mutations and environmental allergen costimulation
- PMID: 29454836
- PMCID: PMC5938139
- DOI: 10.1016/j.jaci.2018.02.003
Mechanism for initiation of food allergy: Dependence on skin barrier mutations and environmental allergen costimulation
Erratum in
-
Correction.J Allergy Clin Immunol. 2018 Jul;142(1):339. doi: 10.1016/j.jaci.2018.05.013. J Allergy Clin Immunol. 2018. PMID: 29980279 No abstract available.
Abstract
Background: Mechanisms for the development of food allergy in neonates are unknown but clearly linked in patient populations to a genetic predisposition to skin barrier defects. Whether skin barrier defects contribute functionally to development of food allergy is unknown.
Objective: The purpose of the study was to determine whether skin barrier mutations, which are primarily heterozygous in patient populations, contribute to the development of food allergy.
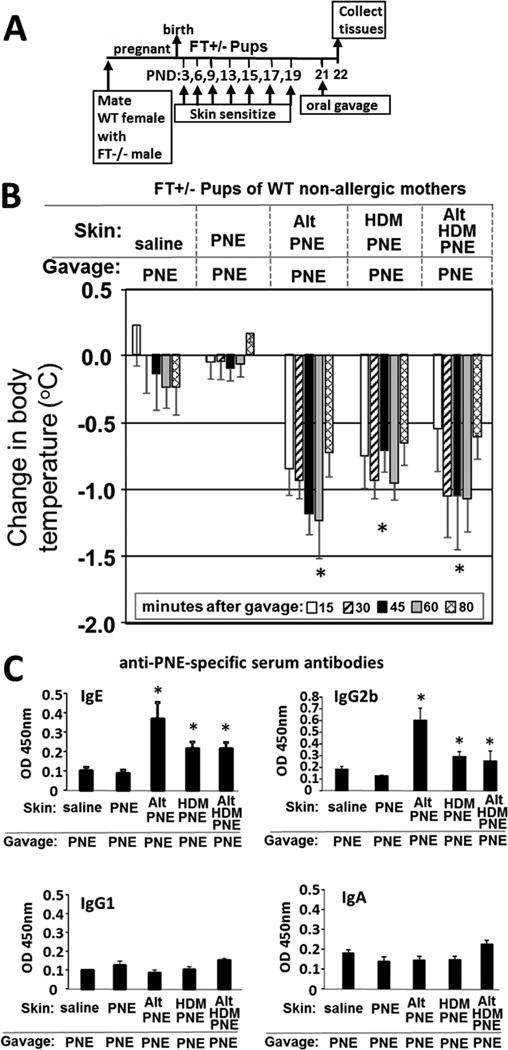
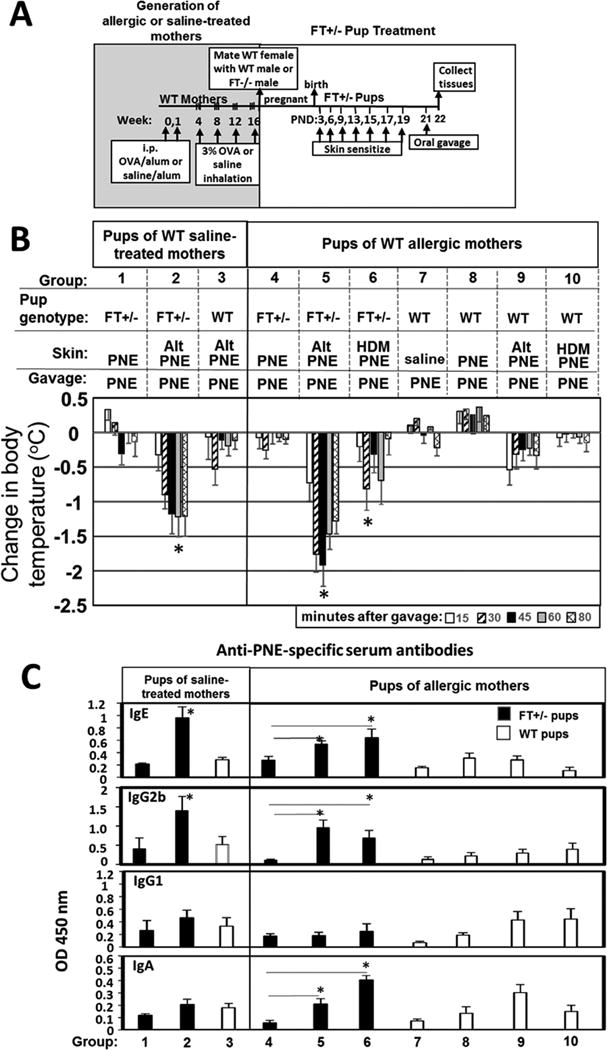
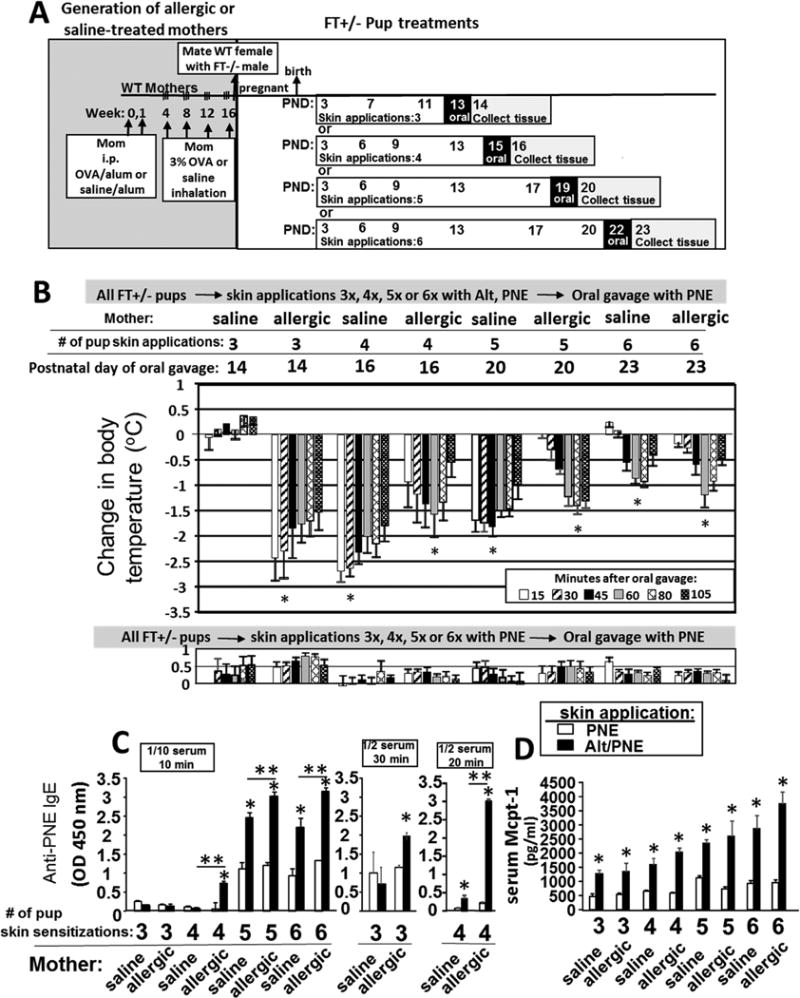
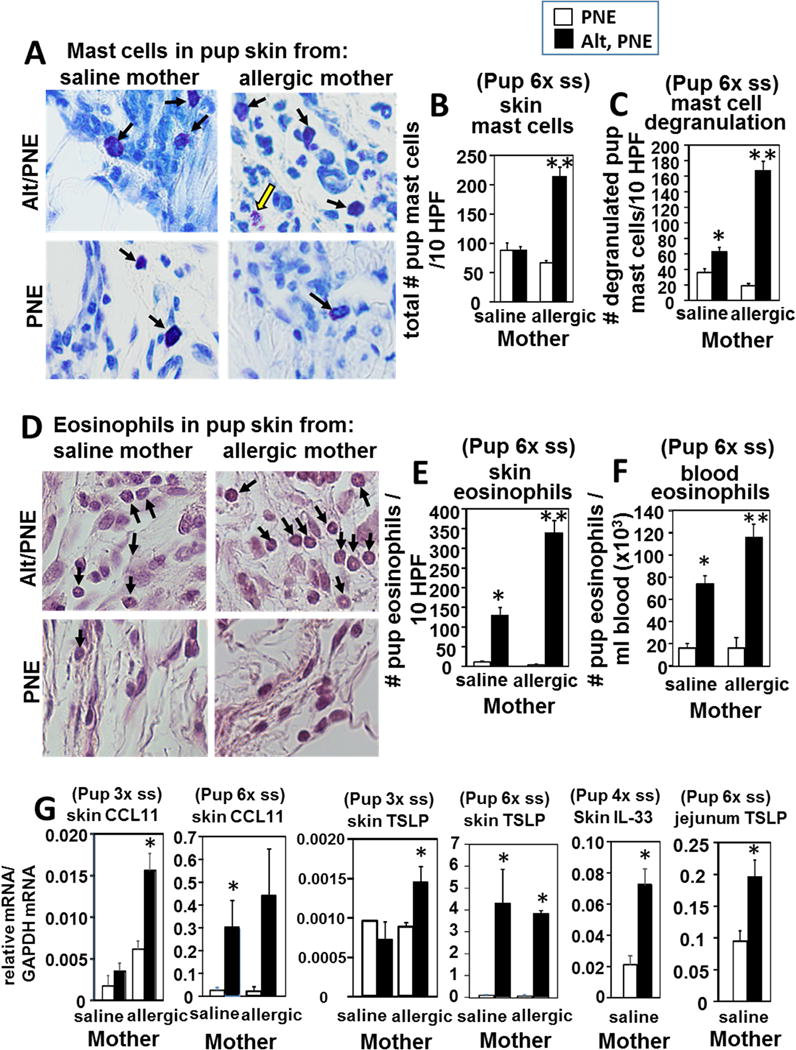
Methods: Mice heterozygous for the filaggrin (Flg)ft and Tmem79ma mutations were skin sensitized with environmental and food allergens. After sensitization, mice received oral challenge with food allergen, and then inflammation, inflammatory mediators, and anaphylaxis were measured.
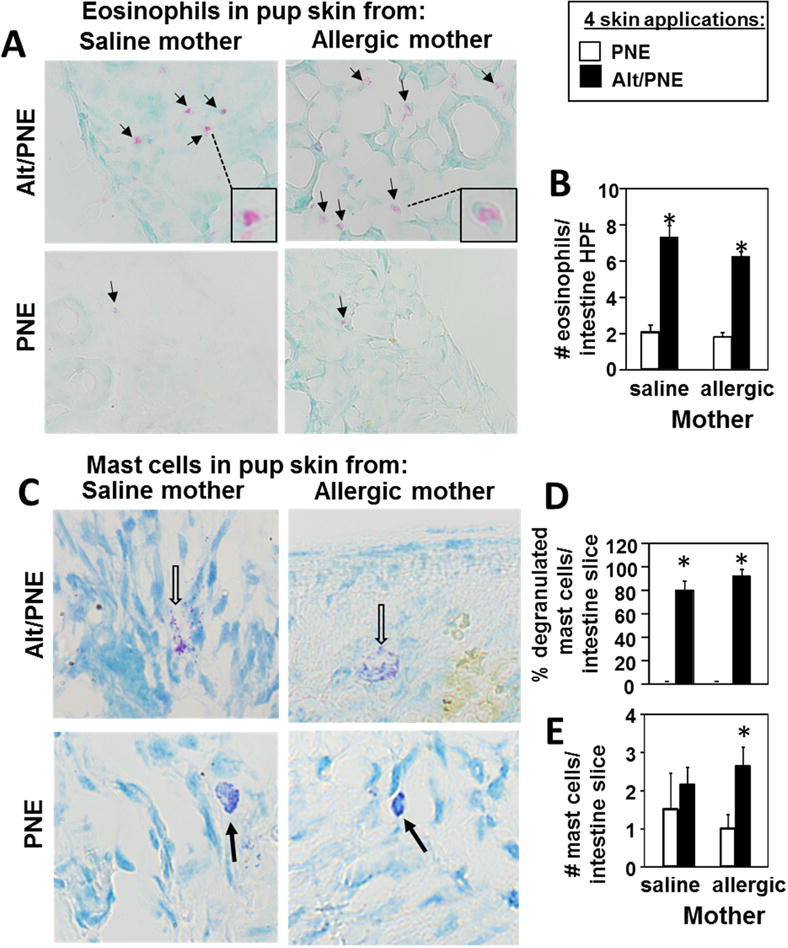
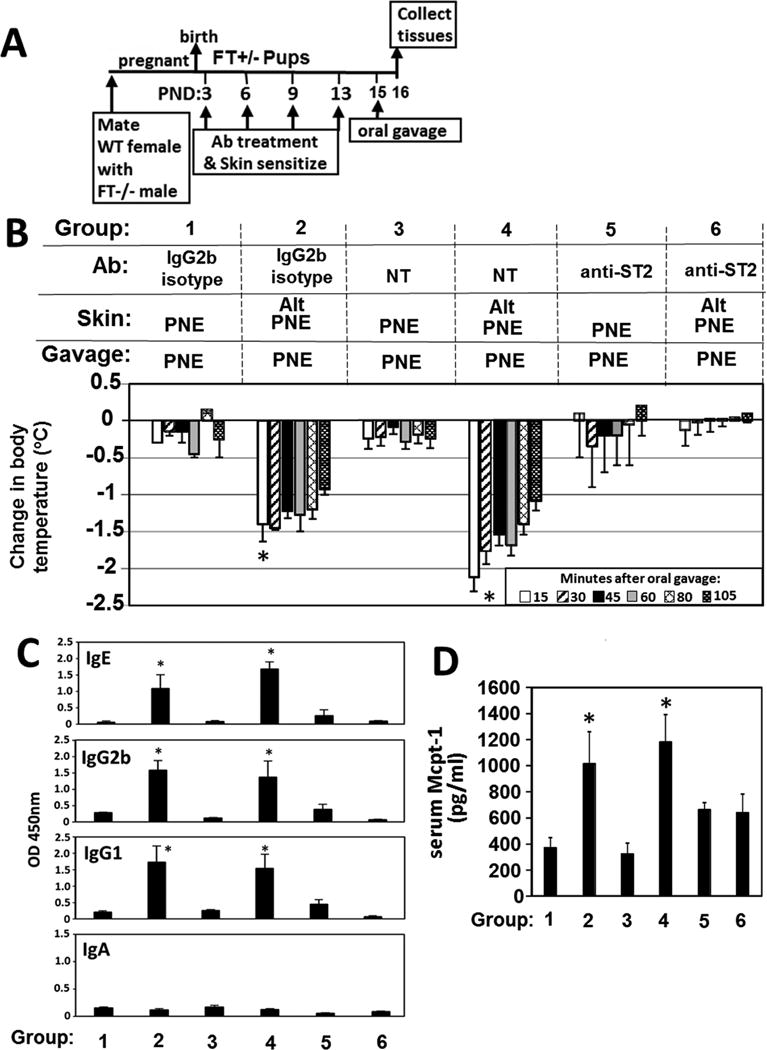
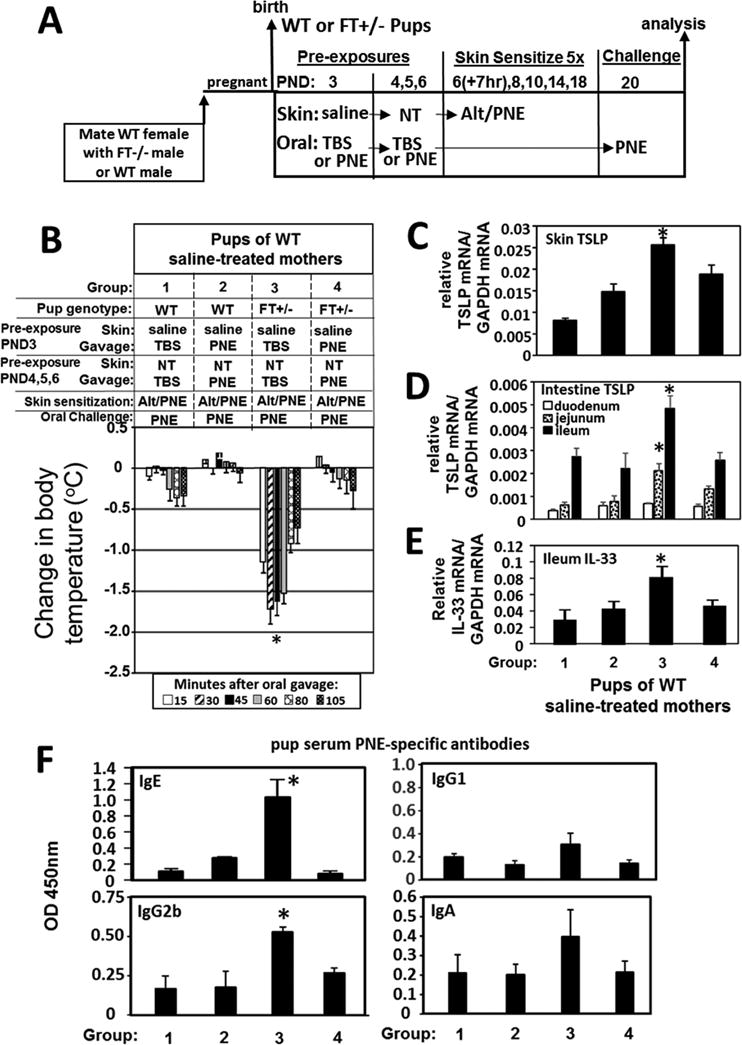
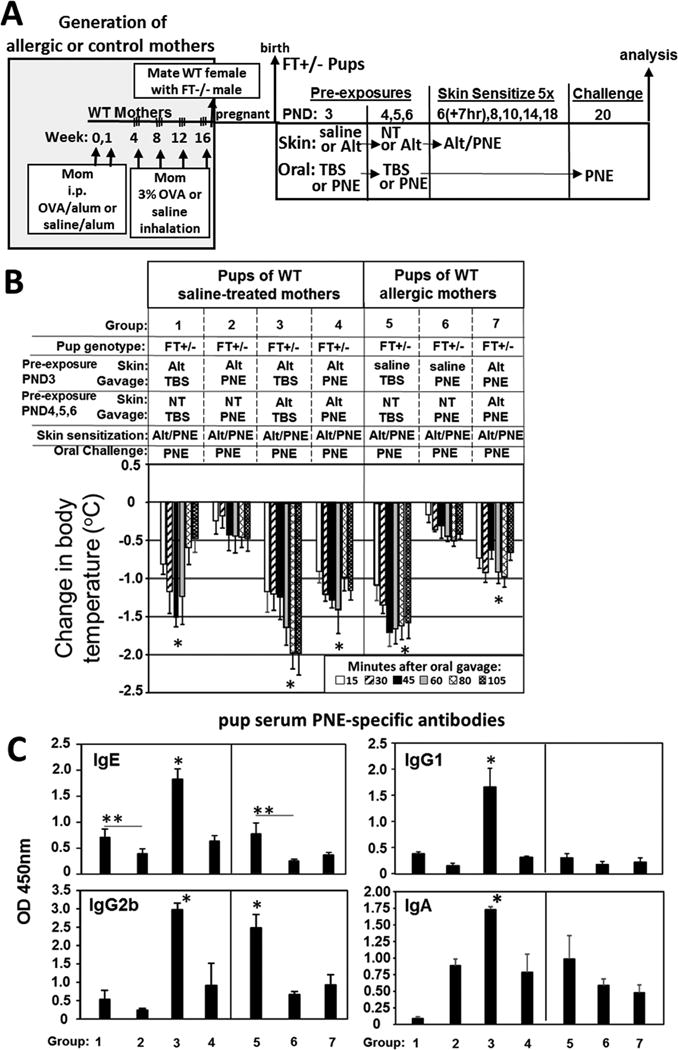
Results: We define development of inflammation, inflammatory mediators, and food allergen-induced anaphylaxis in neonatal mice with skin barrier mutations after brief concurrent cutaneous exposure to food and environmental allergens. Moreover, neonates of allergic mothers have increased responses to suboptimal sensitization with food allergens. Importantly, responses to food allergens by these neonatal mice were dependent on genetic defects in skin barrier function and on exposure to environmental allergens. ST2 blockade during skin sensitization inhibited the development of anaphylaxis, antigen-specific IgE, and inflammatory mediators. Neonatal anaphylactic responses and antigen-specific IgE were also inhibited by oral pre-exposure to food allergen, but interestingly, this was blunted by concurrent pre-exposure of the skin to environmental allergen.
Conclusion: These studies uncover mechanisms for food allergy sensitization and anaphylaxis in neonatal mice that are consistent with features of human early-life exposures and genetics in patients with clinical food allergy and demonstrate that changes in barrier function drive development of anaphylaxis to food allergen.
Keywords: Alternaria alternata; ST2; Sensitization; anaphylaxis; chicken egg ovalbumin; filaggrin; food allergy; house dust mite; mattrin; peanut; skin.
Copyright © 2018 American Academy of Allergy, Asthma & Immunology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- DaVeiga SP. Epidemiology of atopic dermatitis: a review. Allergy Asthma Proc. 2012;33:227–34. - PubMed
-
- Shaker M. New insights into the allergic march. Curr Opin Pediatr. 2014;26:516–20. - PubMed
-
- Bousquet J, Anto J, Auffray C, Akdis M, Cambon-Thomsen A, Keil T, et al. MeDALL (Mechanisms of the Development of ALLergy): an integrated approach from phenotypes to systems medicine. Allergy. 2011;66:596–604. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
