

The Coding Regions of Germline mRNAs Confer Sensitivity to Argonaute Regulation in C. elegans
- PMID: 29456100
- PMCID: PMC5918280
- DOI: 10.1016/j.celrep.2018.02.009
The Coding Regions of Germline mRNAs Confer Sensitivity to Argonaute Regulation in C. elegans
Abstract
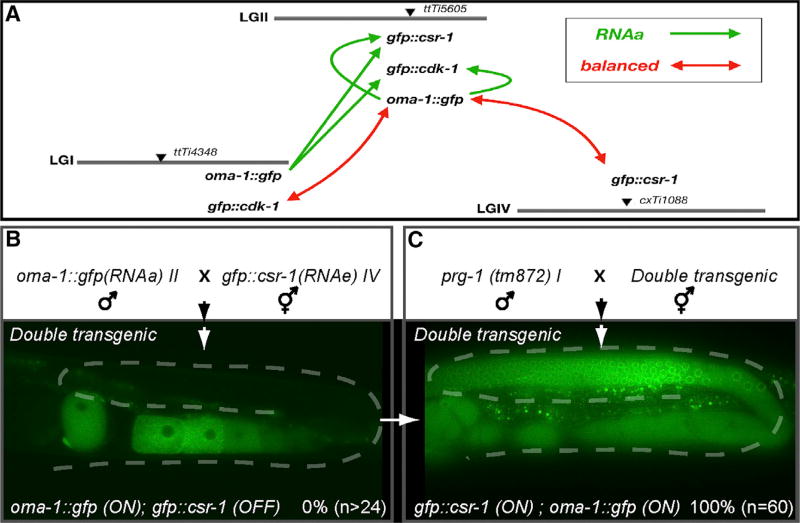
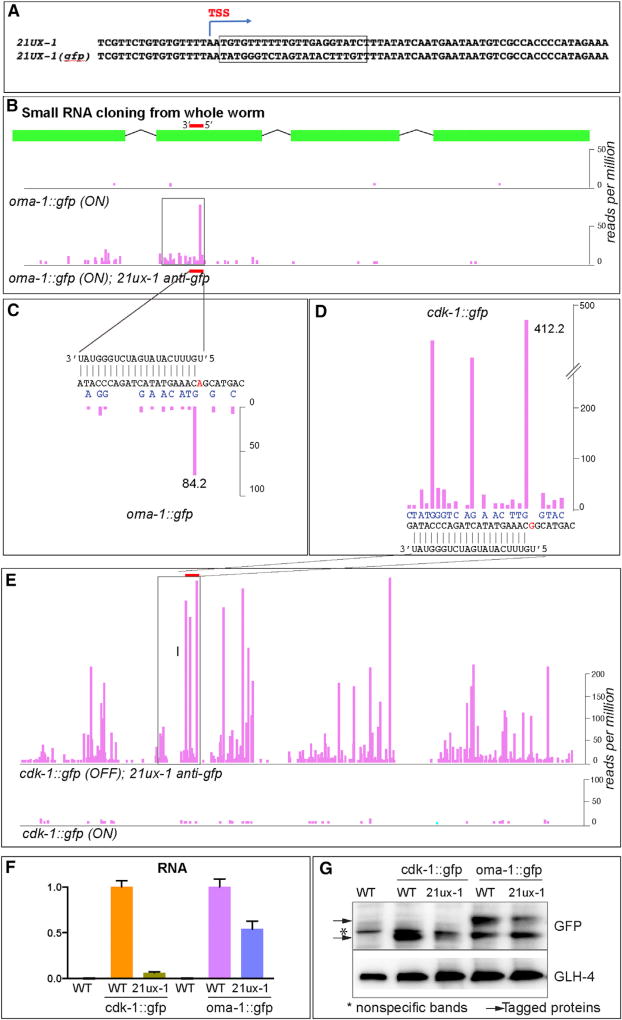
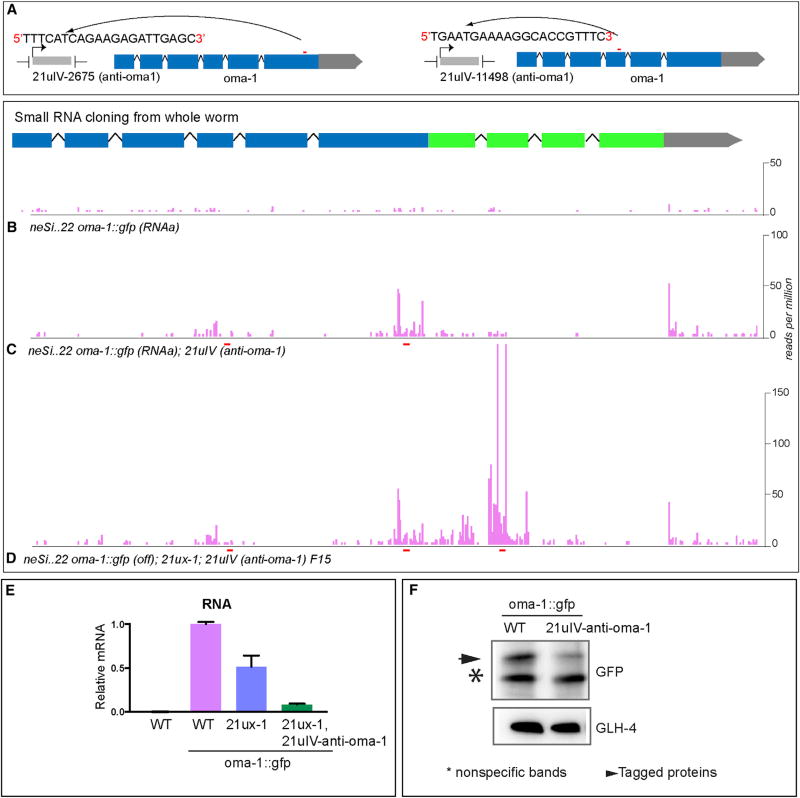
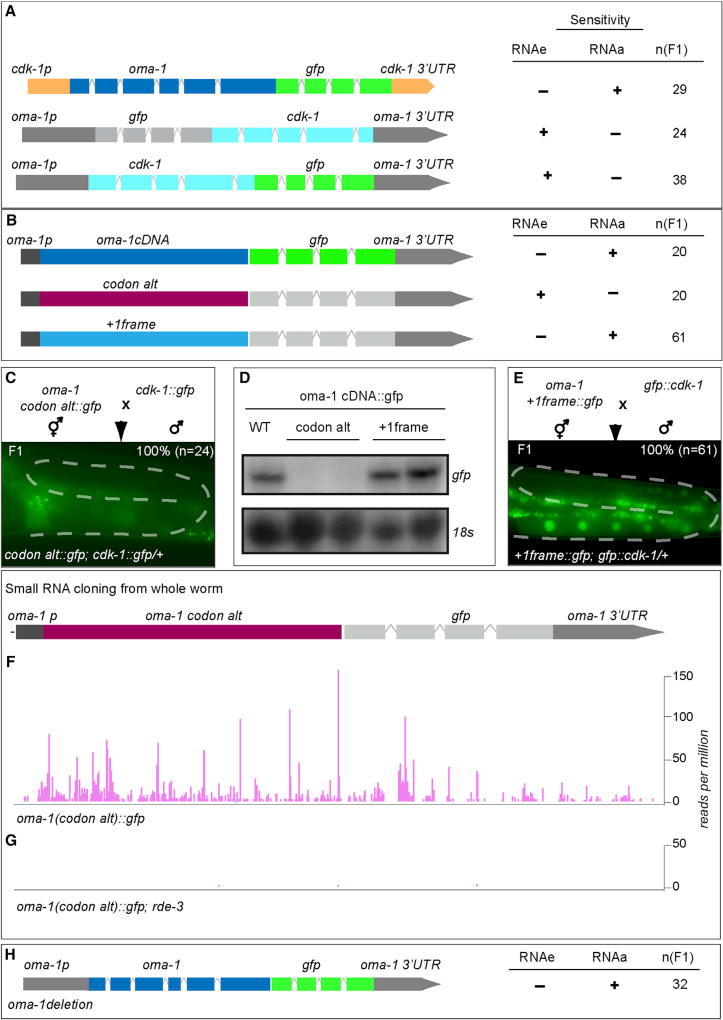
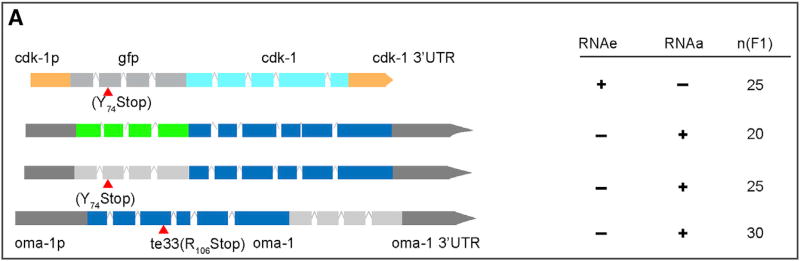
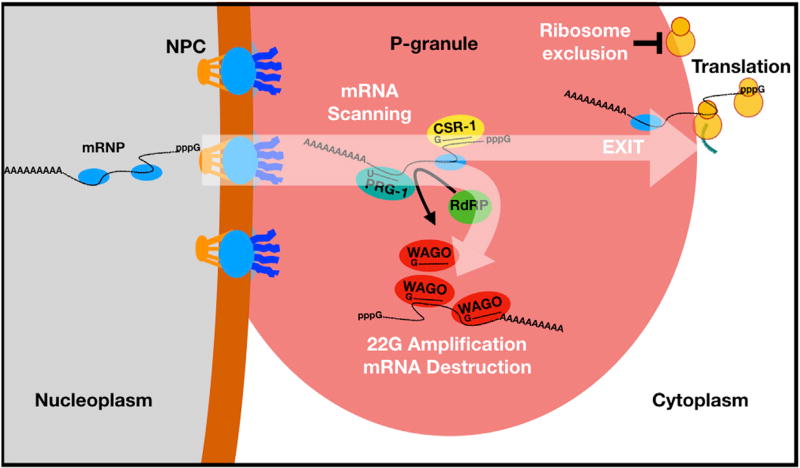
Protein-coding genes undergo a wide array of regulatory interactions with factors that engage non-coding regions. Open reading frames (ORFs), in contrast, are thought to be constrained by coding function, precluding a major role in gene regulation. Here, we explore Piwi-interacting (pi)RNA-mediated transgene silencing in C. elegans and show that marked differences in the sensitivity to piRNA silencing map to the endogenous sequences within transgene ORFs. Artificially increasing piRNA targeting within the ORF of a resistant transgene can lead to a partial yet stable reduction in expression, revealing that piRNAs not only silence but can also "tune" gene expression. Our findings support a model that involves a temporal element to mRNA regulation by germline Argonautes, likely prior to translation, and suggest that piRNAs afford incremental control of germline mRNA expression by targeting the body of the mRNA, including the coding region.
Keywords: Argonautes; C. elegans; P granules; gene expression; nonsense-mediated decay; perinuclear germline nuage; piRNAs.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ, Kuramochi-Miyagawa S, Nakano T, et al. A novel class of small RNAs bind to MILI protein in mouse testes. Nature. 2006;442:203–207. - PubMed
-
- Baker KE, Parker R. Nonsense-mediated mRNA decay: terminating erroneous gene expression. Curr. Opin. Cell Biol. 2004;16:293–299. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
