

Selective Recognition of RNA Substrates by ADAR Deaminase Domains
- PMID: 29457714
- PMCID: PMC5898644
- DOI: 10.1021/acs.biochem.7b01100
Selective Recognition of RNA Substrates by ADAR Deaminase Domains
Abstract
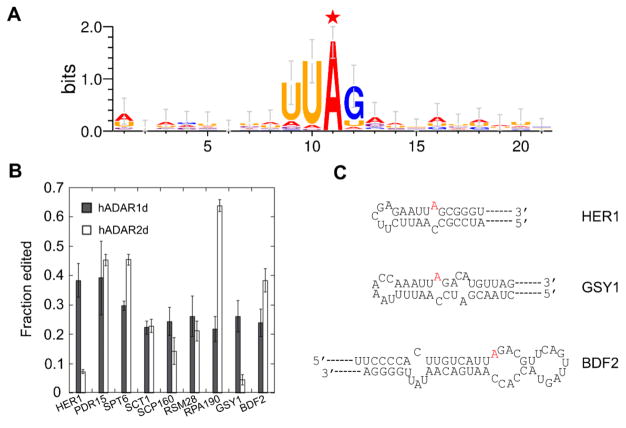
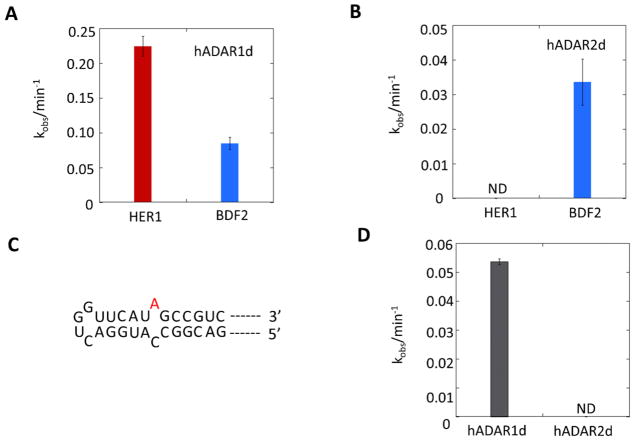
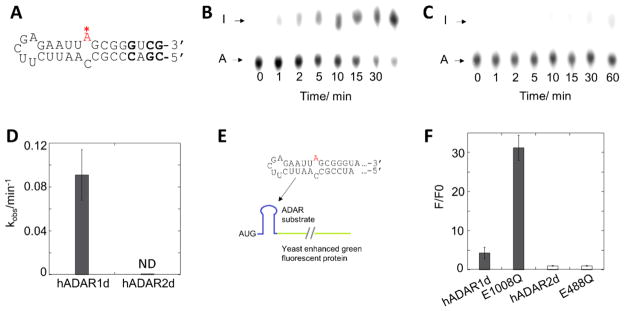
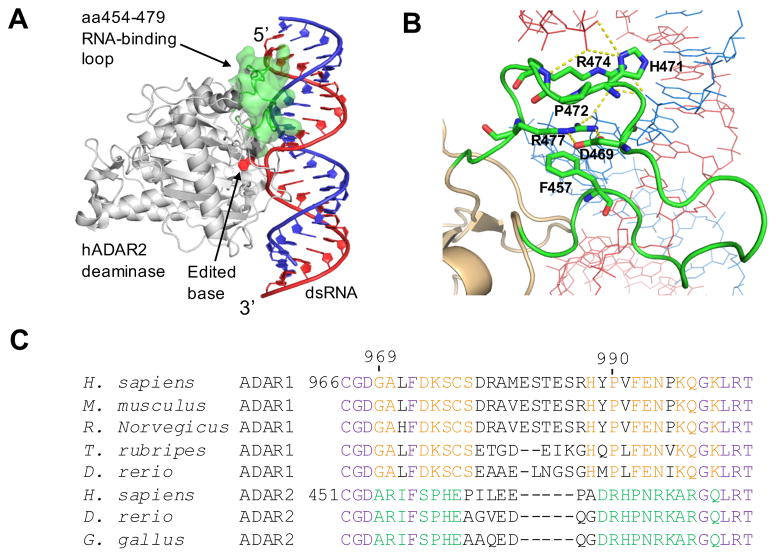
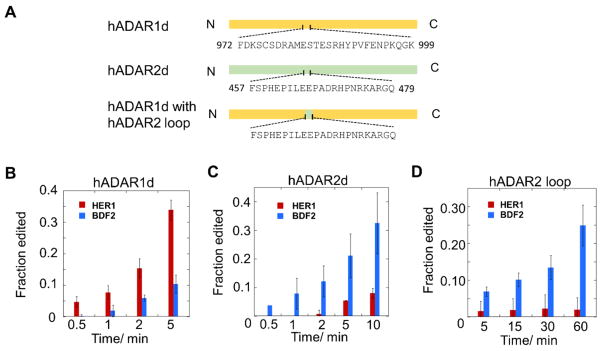
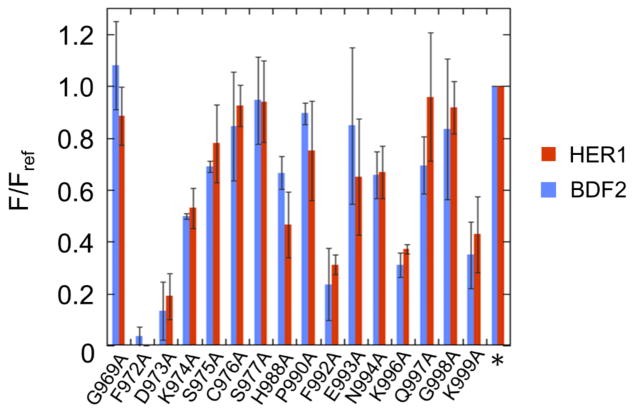
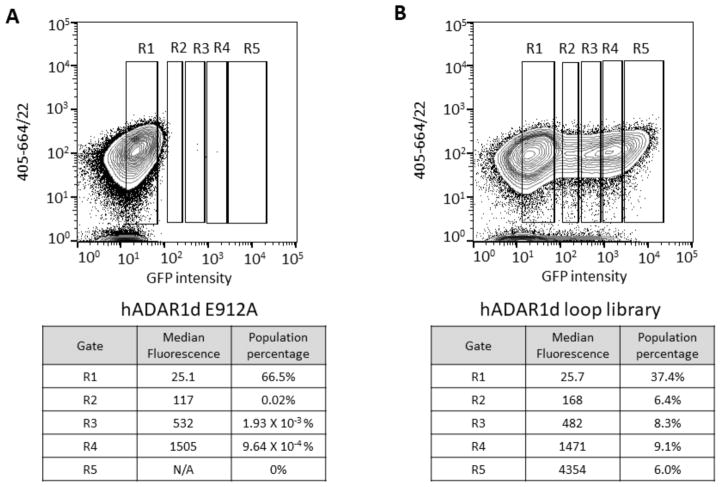
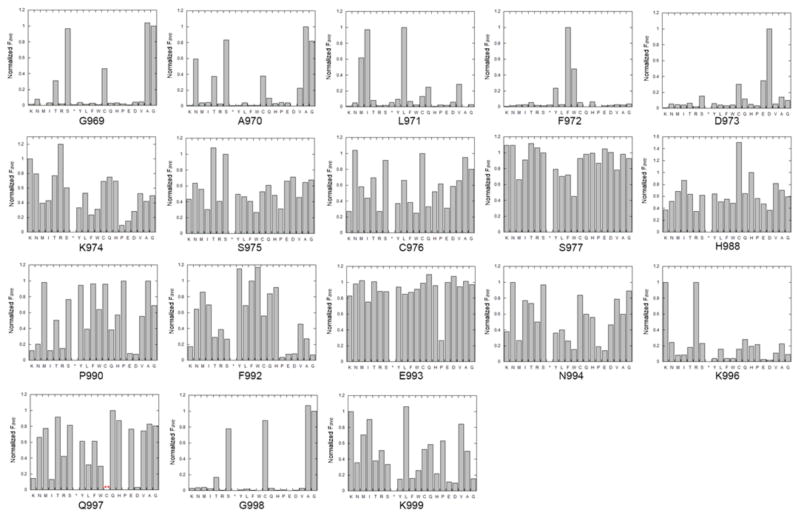
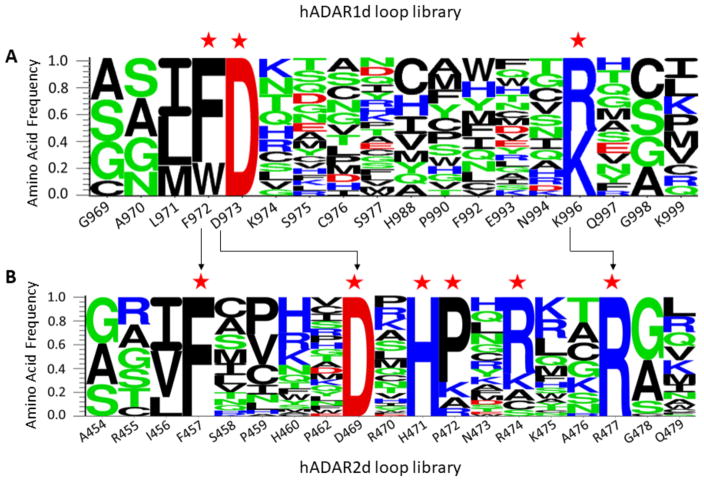
Adenosine deamination is one of the most prevalent post-transcriptional modifications in mRNA and is catalyzed by ADAR1 and ADAR2 in humans. ADAR1 and ADAR2 have different substrate selectivity, which is believed to mainly originate from the proteins' deaminase domains (hADAR1d and hADAR2d, respectively). RNA-seq of the Saccharomyces cerevisiae transcriptome subjected to ADAR-catalyzed RNA editing identified substrates with common secondary structure features preferentially edited by hADAR1d over hADAR2d. The relatively small size and efficient reaction of one of these substrates suggested it could be useful for further study of the hADAR1d reaction. Indeed, a short hairpin stem from the S. cerevisiae HER1 mRNA was efficiently deaminated by hADAR1d and used to generate an hADAR1d-specific fluorescent reporter of editing activity. Using substrates preferred by either hADAR1d or hADAR2d in vitro, we found that a chimeric protein bearing an RNA-binding loop from hADAR2d grafted onto hADAR1d showed ADAR2-like selectivity. Finally, a high-throughput mutagenesis analysis (Sat-FACS-Seq) of conserved residues in an RNA-binding loop of hADAR1d revealed essential amino acids for function, advancing our understanding of RNA recognition by this domain.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Li X, Xiong X, Wang K, Wang L, Shu X, Ma S, Yi C. Transcriptome-wide mapping reveals reversible and dynamic N1-methyladenosine methylome. Nat Chem Biol. 2016;12:311–316. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
