

Dynamics of promoter bivalency and RNAP II pausing in mouse stem and differentiated cells
- PMID: 29458328
- PMCID: PMC5819258
- DOI: 10.1186/s12861-018-0163-7
Dynamics of promoter bivalency and RNAP II pausing in mouse stem and differentiated cells
Abstract
Background: Mammalian embryonic stem cells display a unique epigenetic and transcriptional state to facilitate pluripotency by maintaining lineage-specification genes in a poised state. Two epigenetic and transcription processes involved in maintaining poised state are bivalent chromatin, characterized by the simultaneous presence of activating and repressive histone methylation marks, and RNA polymerase II (RNAPII) promoter proximal pausing. However, the dynamics of histone modifications and RNAPII at promoters in diverse cellular contexts remains underexplored.
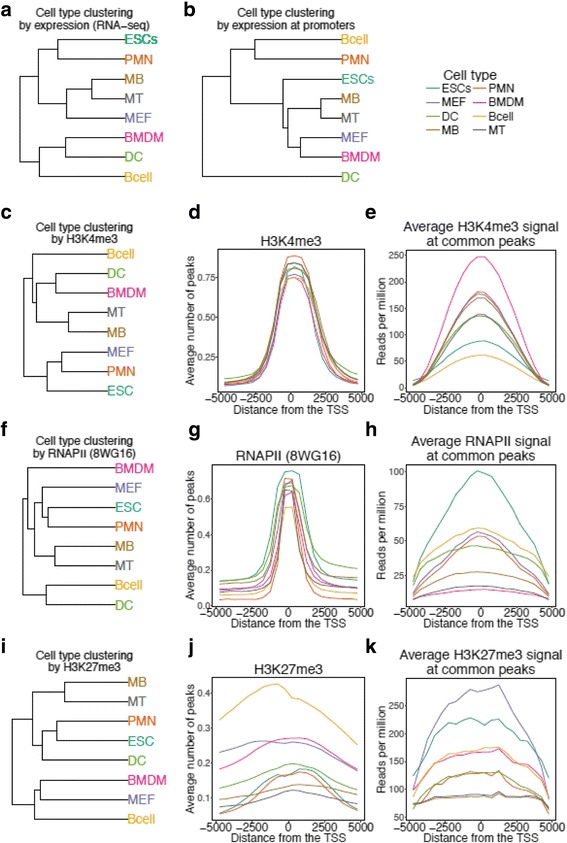
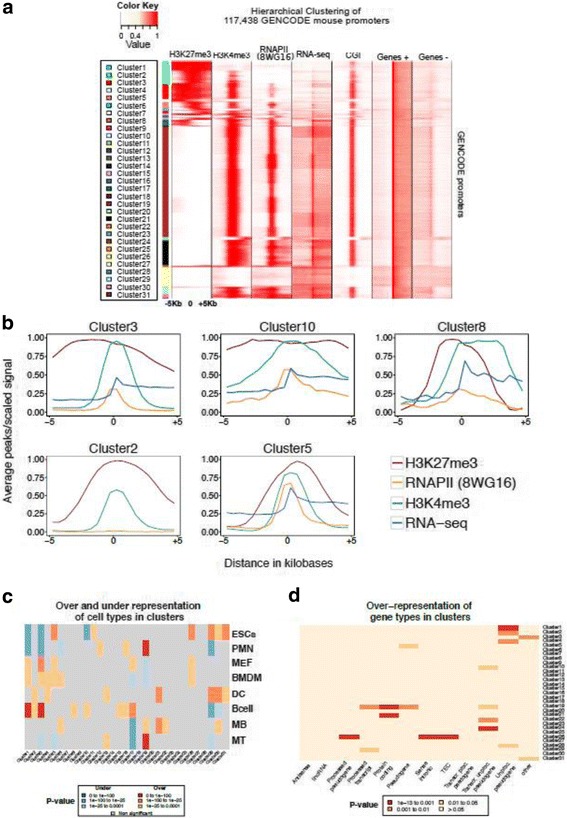
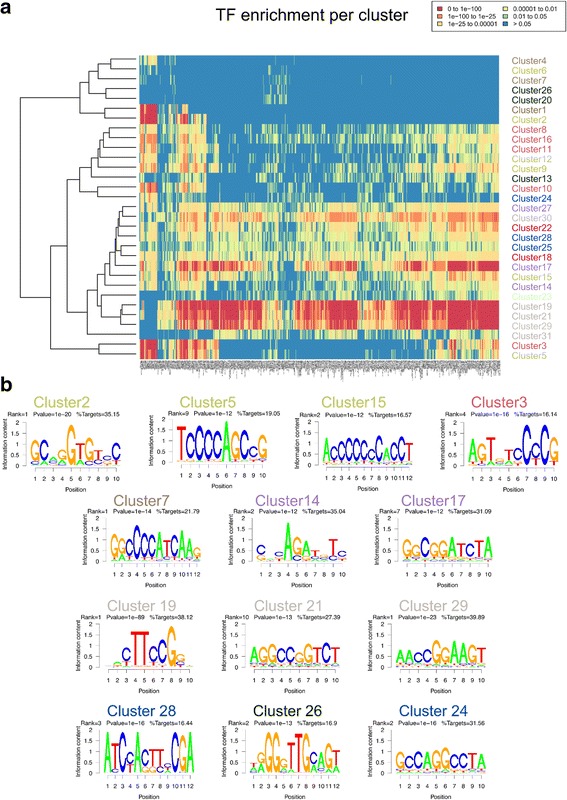
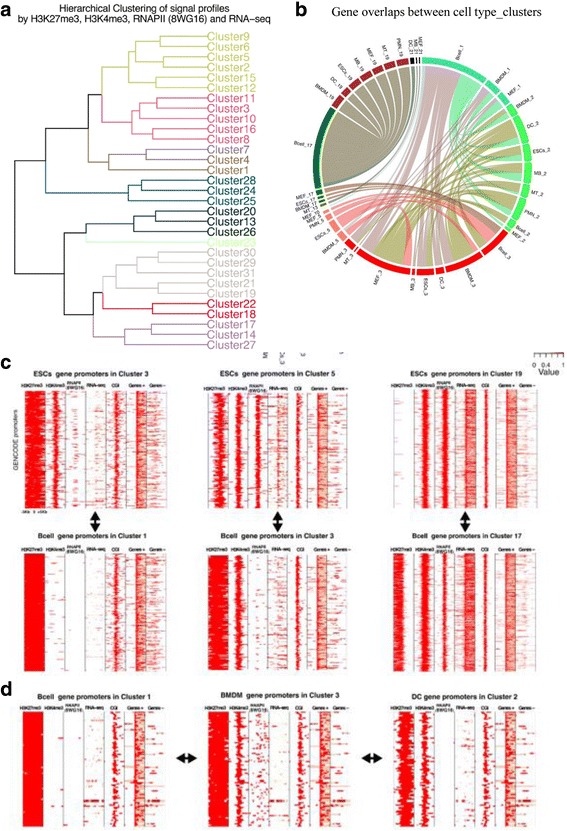
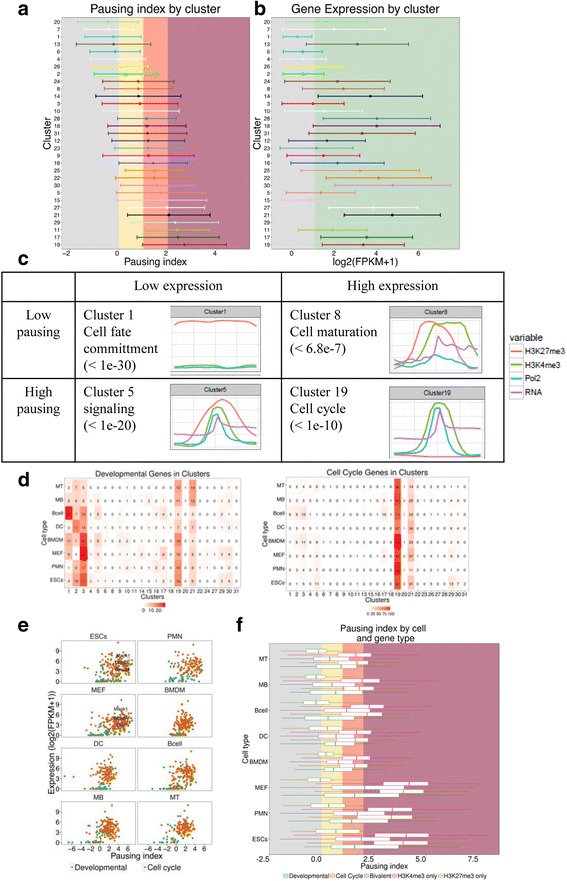
Results: We collected genome wide data for bivalent chromatin marks H3K4me3 and H3K27me3, and RNAPII (8WG16) occupancy together with expression profiling in eight different cell types, including ESCs, in mouse. The epigenetic and transcription profiles at promoters grouped in over thirty clusters with distinct functional identities and transcription control.
Conclusion: The clustering analysis identified distinct bivalent clusters where genes in one cluster retained bivalency across cell types while in the other were mostly cell type specific, but neither showed a high RNAPII pausing. We noted that RNAPII pausing is more associated with active genes than bivalent genes in a cell type, and was globally reduced in differentiated cell types compared to multipotent.
Keywords: Bivalent; Chromatin; ES cells; Gene expression; Pausing; RNA pol II.
Conflict of interest statement
Ethics approval and consent to participate
Not Applicable
Consent for publication
Not Applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Reik W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nat Publ Group. 2007;447:425–432. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
