

Neuromodulation via the Cerebrospinal Fluid: Insights from Recent in Vitro Studies
- PMID: 29459822
- PMCID: PMC5807333
- DOI: 10.3389/fncir.2018.00005
Neuromodulation via the Cerebrospinal Fluid: Insights from Recent in Vitro Studies
Abstract
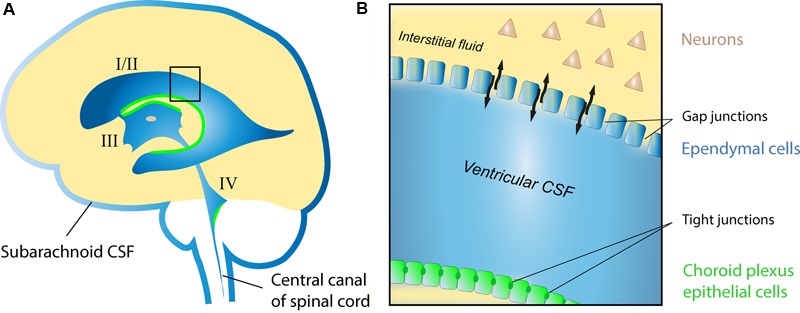
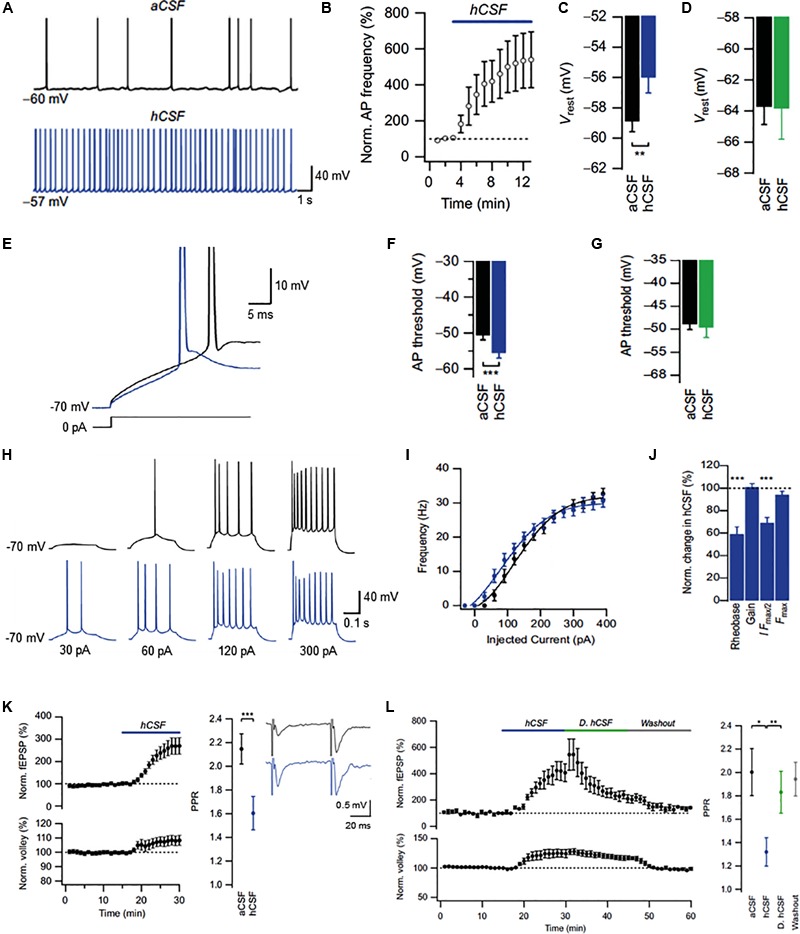
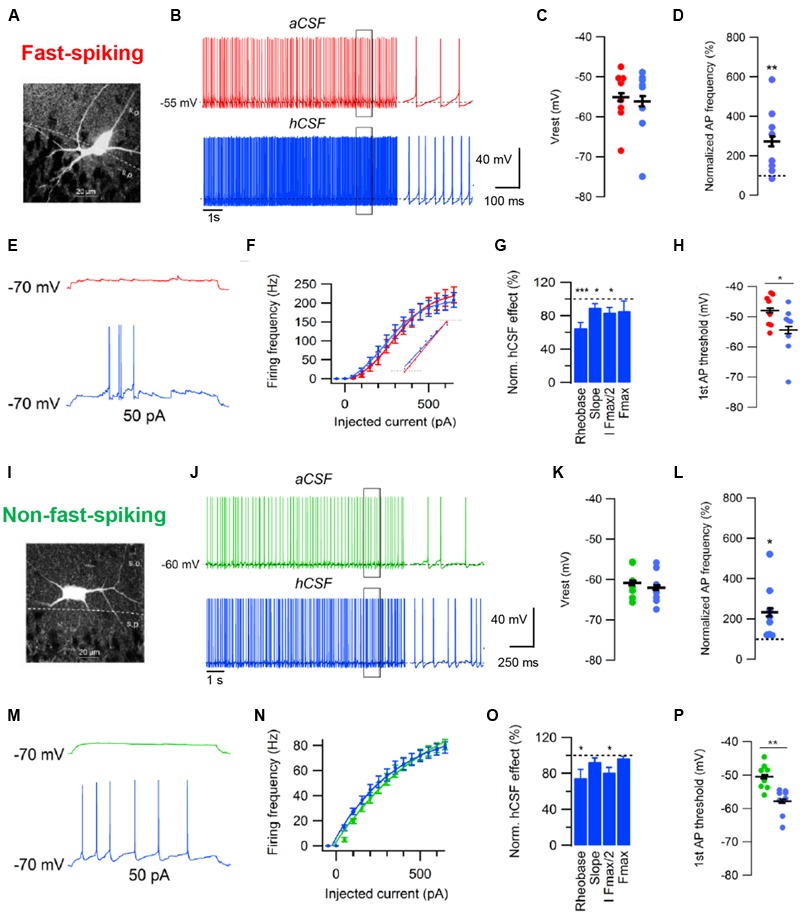
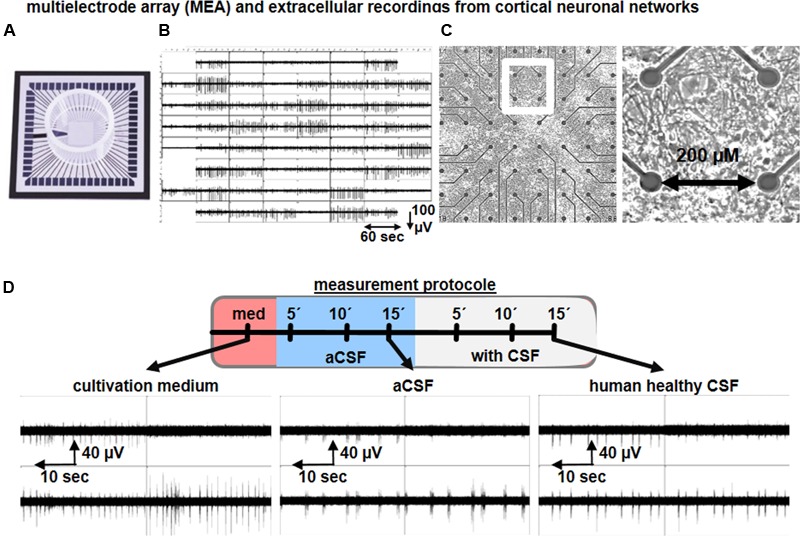
The cerebrospinal fluid (CSF) occupies the brain's ventricles and subarachnoid space and, together with the interstitial fluid (ISF), forms a continuous fluidic network that bathes all cells of the central nervous system (CNS). As such, the CSF is well positioned to actively distribute neuromodulators to neural circuits in vivo via volume transmission. Recent in vitro experimental work in brain slices and neuronal cultures has shown that human CSF indeed contains neuromodulators that strongly influence neuronal activity. Here we briefly summarize these new findings and discuss their potential relevance to neural circuits in health and disease.
Keywords: cerebrospinal fluid; neural circuit; neuromodulation.
Figures
References
-
- Agnati L. F., Fuxe K., Zoli M., Ozini I., Toffano G., Ferraguti F. (1986). A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta Physiol. Scand. 128 201–207. 10.1111/j.1748-1716.1986.tb07967.x - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials