

Evolving artificial metalloenzymes via random mutagenesis
- PMID: 29461523
- PMCID: PMC5891097
- DOI: 10.1038/nchem.2927
Evolving artificial metalloenzymes via random mutagenesis
Abstract
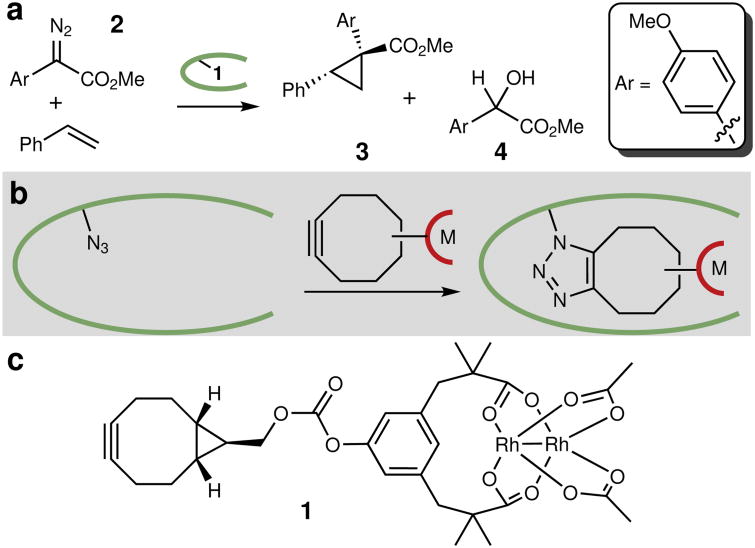
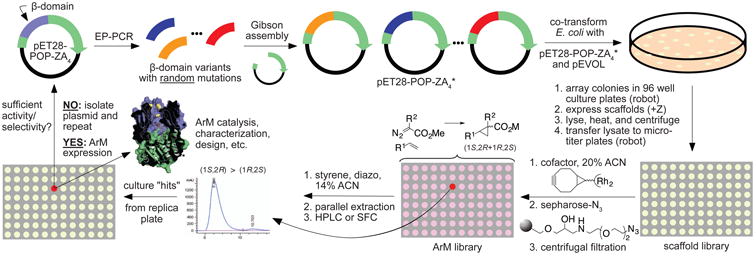
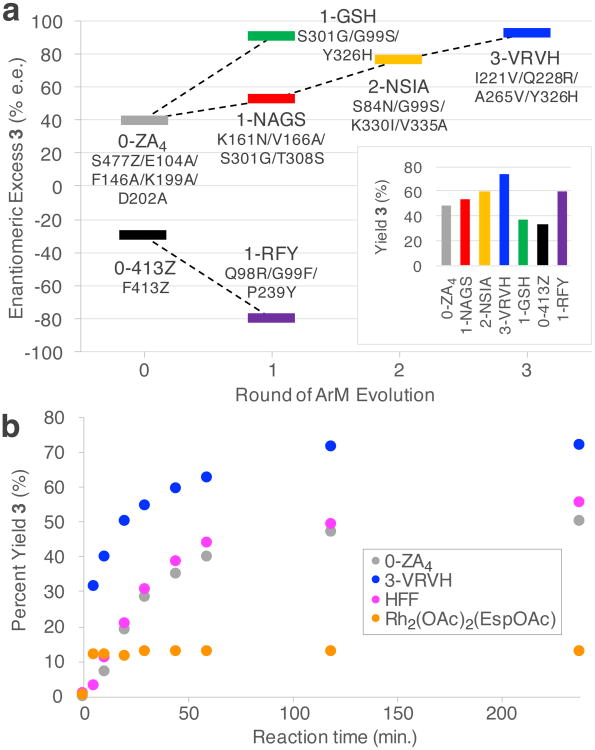
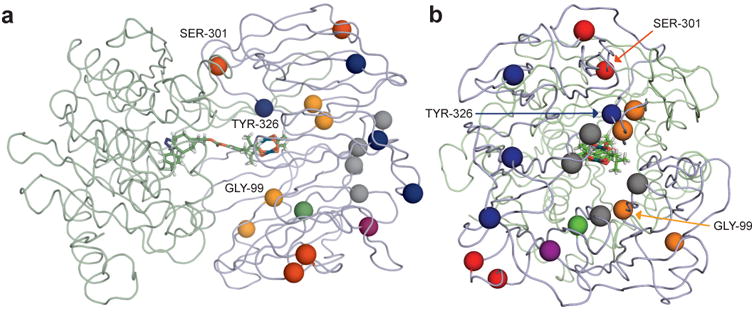
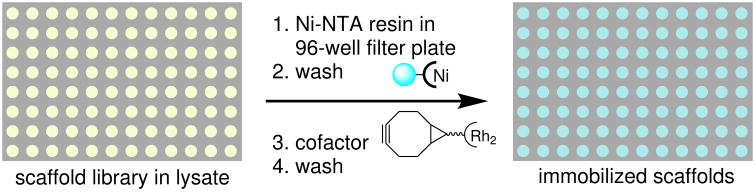
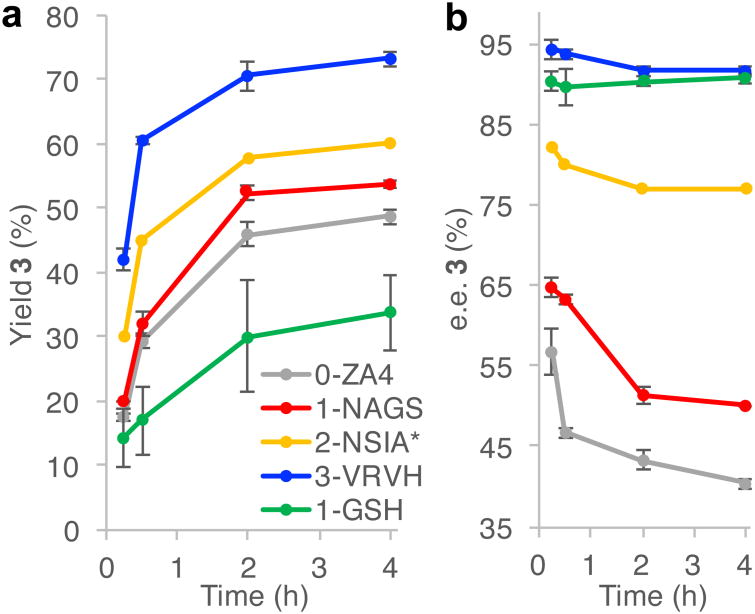
Random mutagenesis has the potential to optimize the efficiency and selectivity of protein catalysts without requiring detailed knowledge of protein structure; however, introducing synthetic metal cofactors complicates the expression and screening of enzyme libraries, and activity arising from free cofactor must be eliminated. Here we report an efficient platform to create and screen libraries of artificial metalloenzymes (ArMs) via random mutagenesis, which we use to evolve highly selective dirhodium cyclopropanases. Error-prone PCR and combinatorial codon mutagenesis enabled multiplexed analysis of random mutations, including at sites distal to the putative ArM active site that are difficult to identify using targeted mutagenesis approaches. Variants that exhibited significantly improved selectivity for each of the cyclopropane product enantiomers were identified, and higher activity than previously reported ArM cyclopropanases obtained via targeted mutagenesis was also observed. This improved selectivity carried over to other dirhodium-catalysed transformations, including N-H, S-H and Si-H insertion, demonstrating that ArMs evolved for one reaction can serve as starting points to evolve catalysts for others.
Figures
References
-
- Mahatthananchai J, Dumas AM, Bode JW. Catalytic Selective Synthesis. Angew Chem Int Ed. 2012;51:10954–10990. - PubMed
-
- Bornscheuer UT, et al. Engineering the Third Wave of Biocatalysis. Nature. 2012;485:185–194. - PubMed
-
- Lu Y, Berry S, Pfister T. Engineering novel metalloproteins: Design of metal-binding sites into native protein scaffolds. Chem Rev. 2001;101:3047–3080. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
