

Direct observation of the influence of cardiolipin and antibiotics on lipid II binding to MurJ
- PMID: 29461535
- PMCID: PMC5912511
- DOI: 10.1038/nchem.2919
Direct observation of the influence of cardiolipin and antibiotics on lipid II binding to MurJ
Abstract
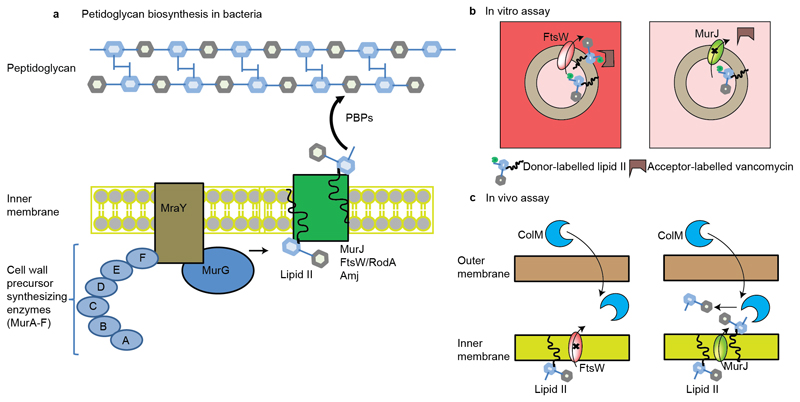
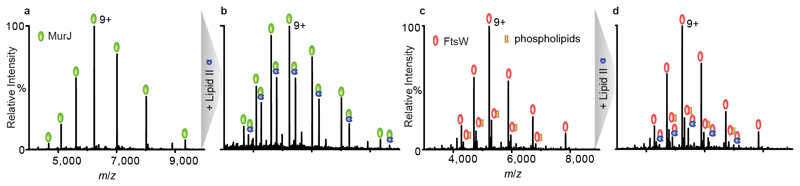
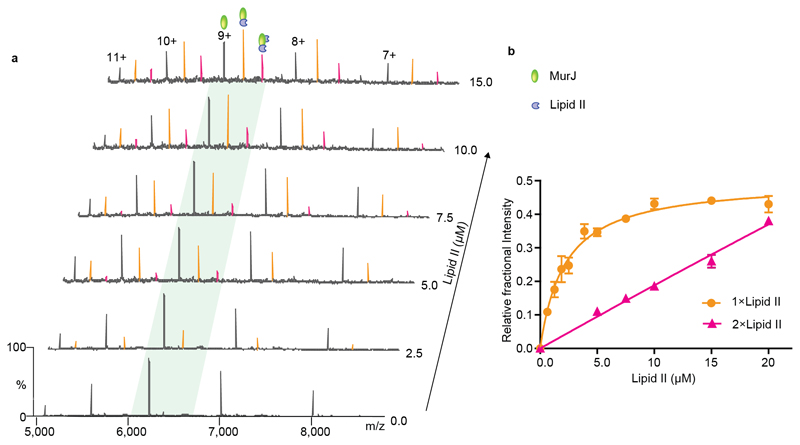
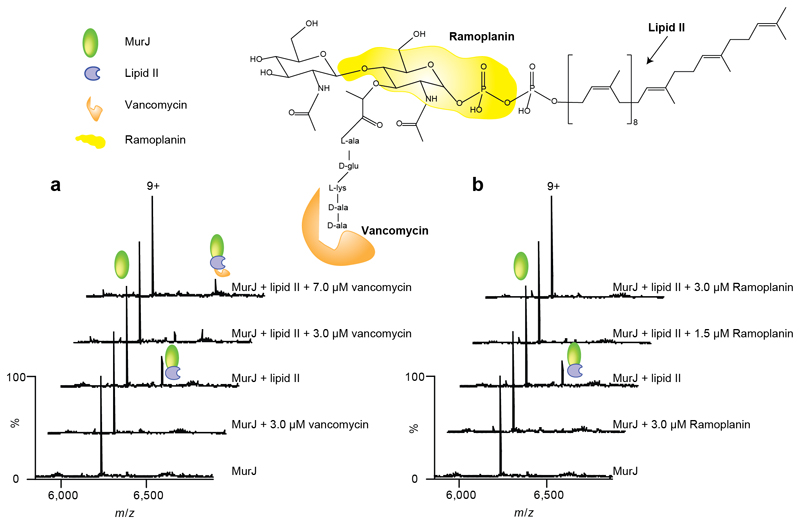
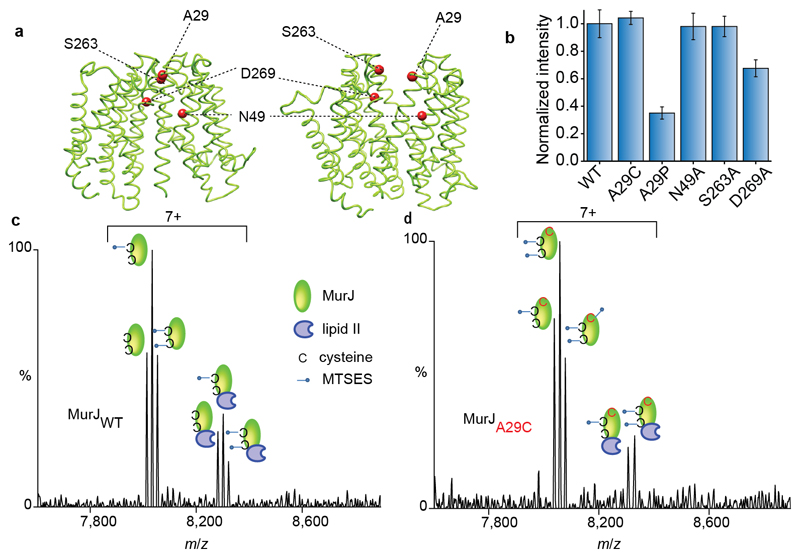
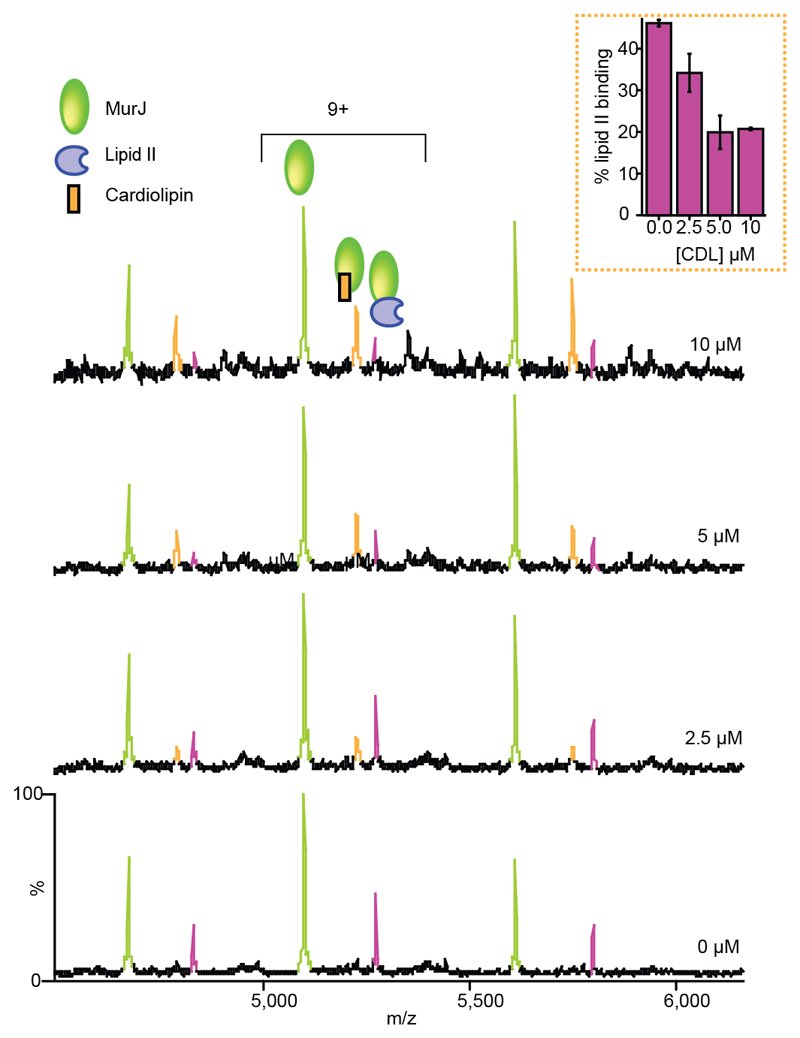
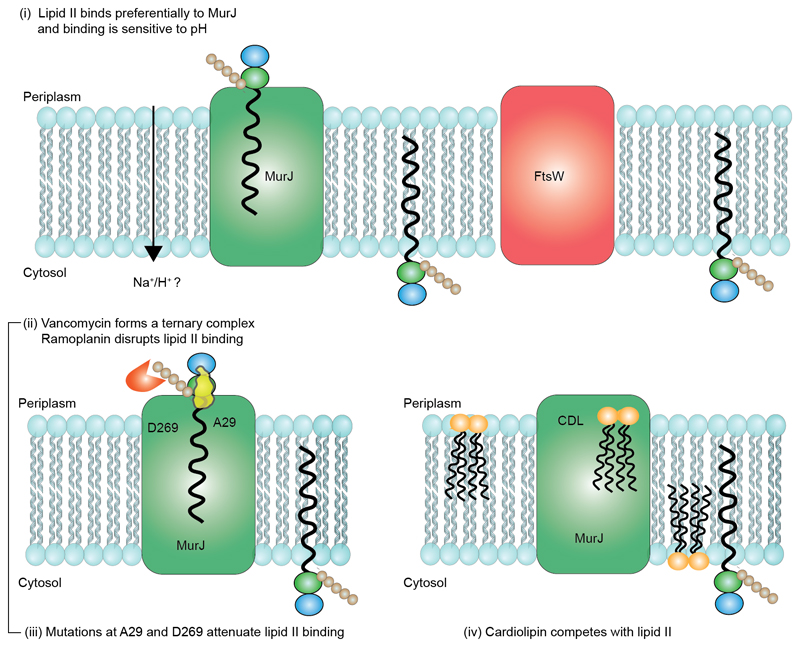
Translocation of lipid II across the cytoplasmic membrane is essential in peptidoglycan biogenesis. Although most steps are understood, identifying the lipid II flippase has yielded conflicting results, and the lipid II binding properties of two candidate flippases-MurJ and FtsW-remain largely unknown. Here we apply native mass spectrometry to both proteins and characterize lipid II binding. We observed lower levels of lipid II binding to FtsW compared to MurJ, consistent with MurJ having a higher affinity. Site-directed mutagenesis of MurJ suggests that mutations at A29 and D269 attenuate lipid II binding to MurJ, whereas chemical modification of A29 eliminates binding. The antibiotic ramoplanin dissociates lipid II from MurJ, whereas vancomycin binds to form a stable complex with MurJ:lipid II. Furthermore, we reveal cardiolipins associate with MurJ but not FtsW, and exogenous cardiolipins reduce lipid II binding to MurJ. These observations provide insights into determinants of lipid II binding to MurJ and suggest roles for endogenous lipids in regulating substrate binding.
Figures
References
-
- de Kruijff B, van Dam V, Breukink E. Lipid II: a central component in bacterial cell wall synthesis and a target for antibiotics. Prostaglandins Leukot Essent Fatty Acids. 2008;79:117–121. - PubMed
-
- Schneider T, Sahl H-GG. An oldie but a goodie - cell wall biosynthesis as antibiotic target pathway. Int J Med Microbiol. 2010;300:161–169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
