

The INO80 chromatin remodeler sustains metabolic stability by promoting TOR signaling and regulating histone acetylation
- PMID: 29462149
- PMCID: PMC5834206
- DOI: 10.1371/journal.pgen.1007216
The INO80 chromatin remodeler sustains metabolic stability by promoting TOR signaling and regulating histone acetylation
Abstract
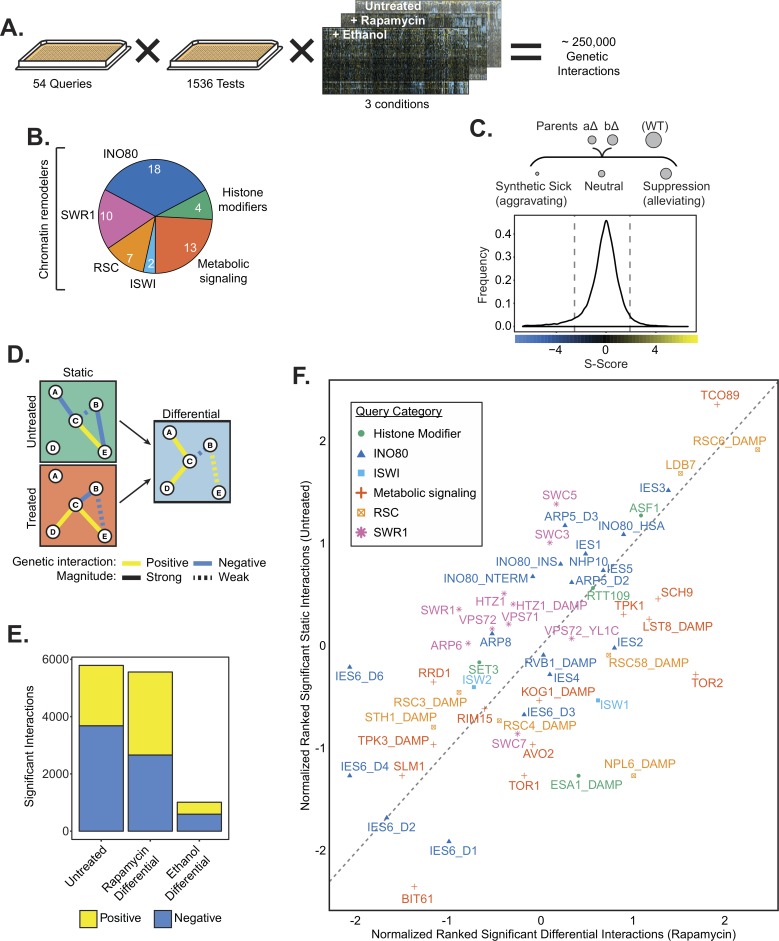
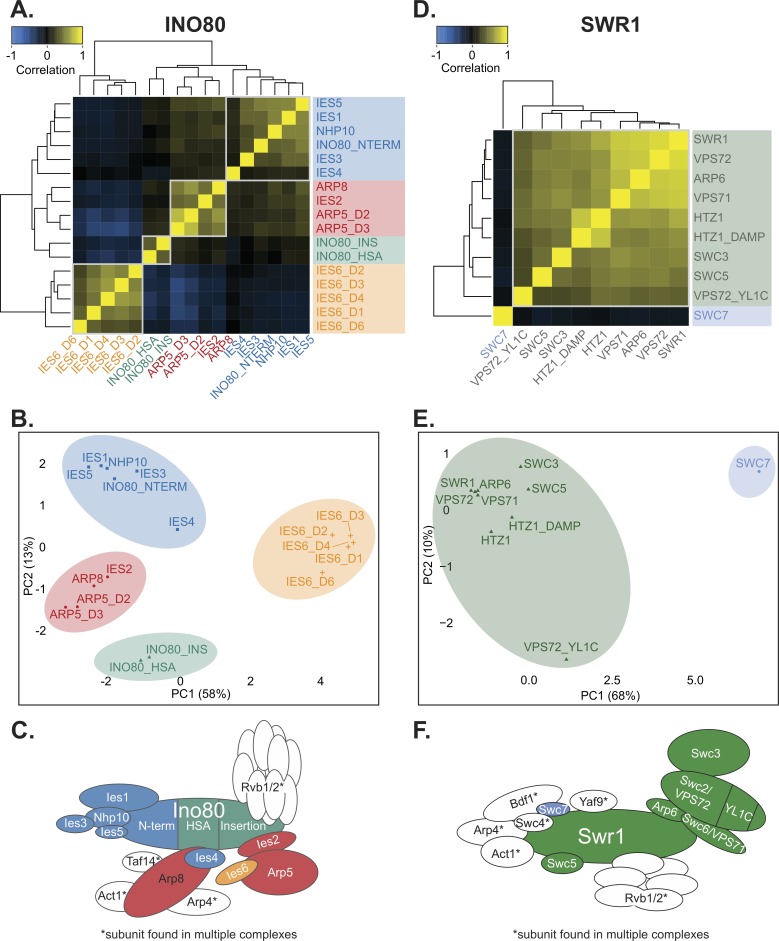
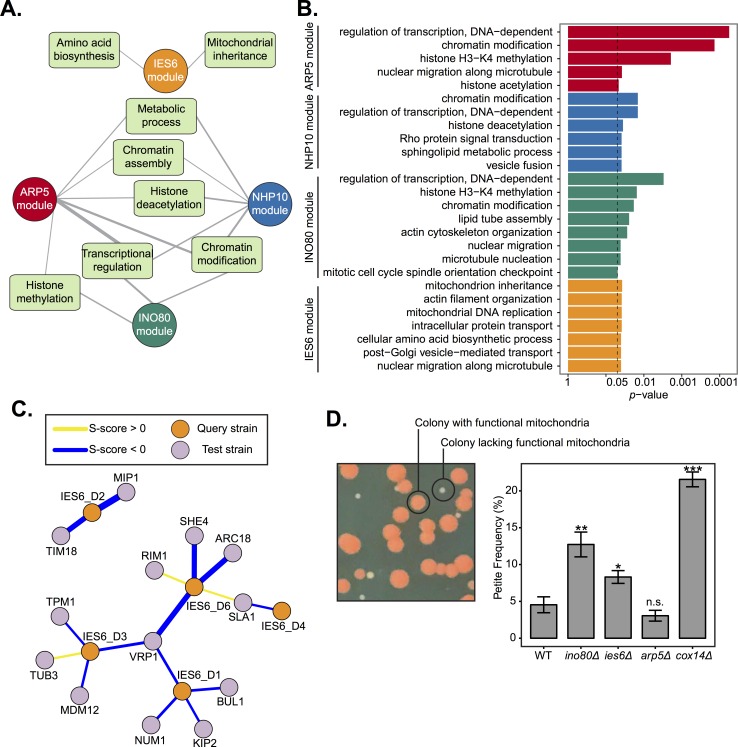
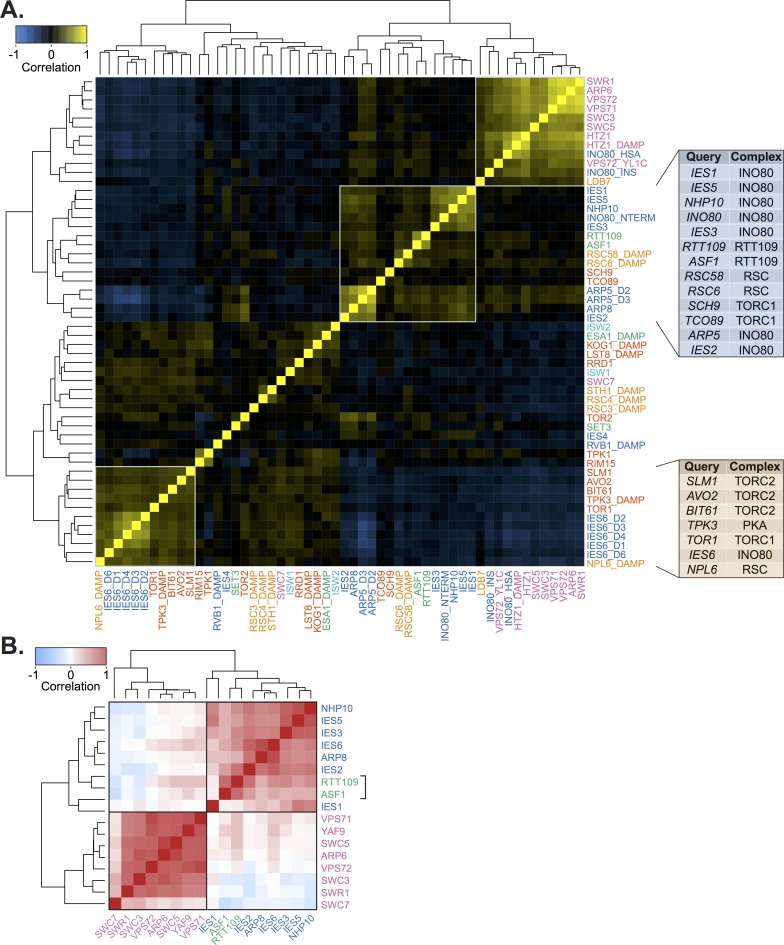
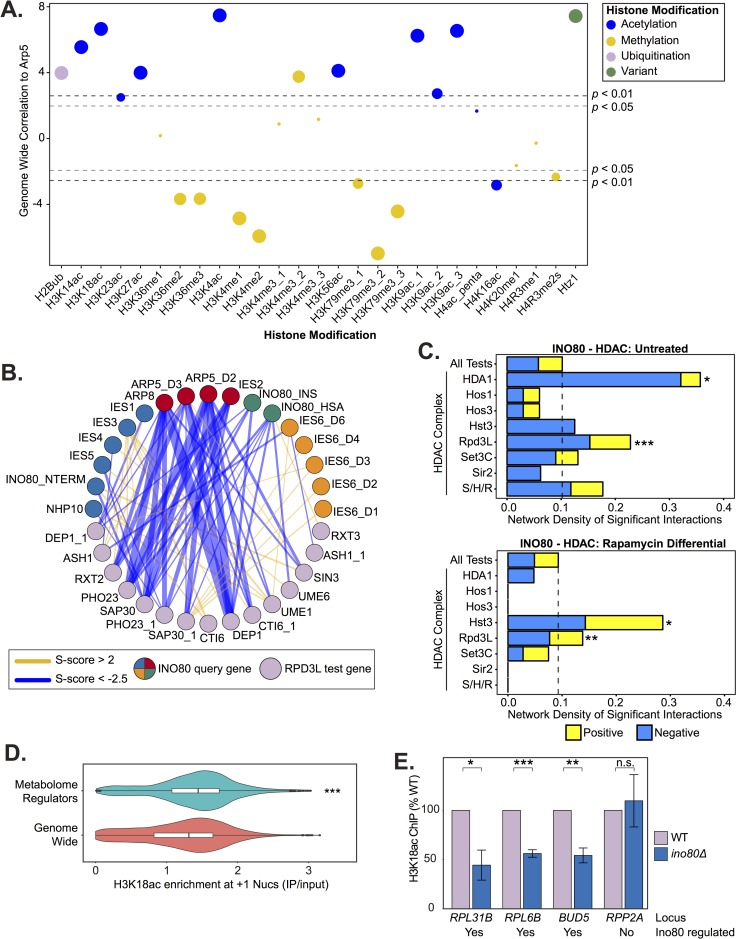
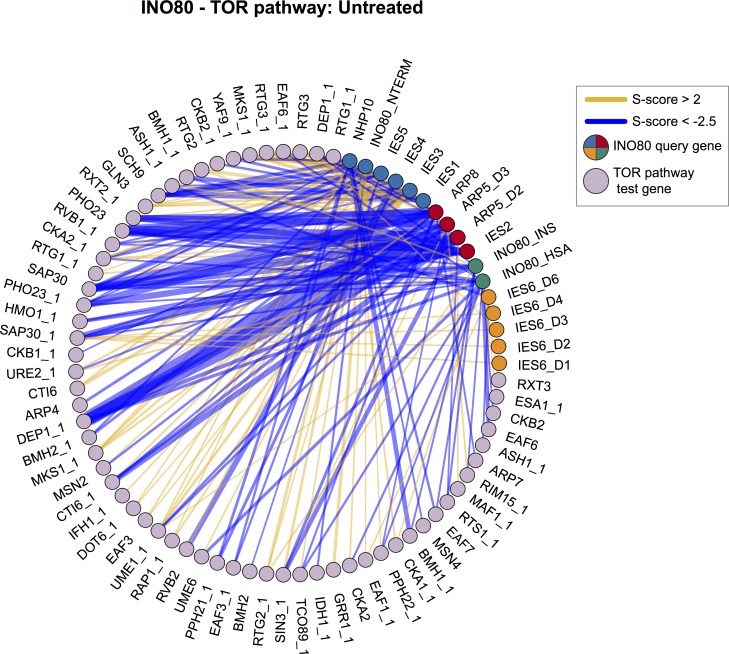
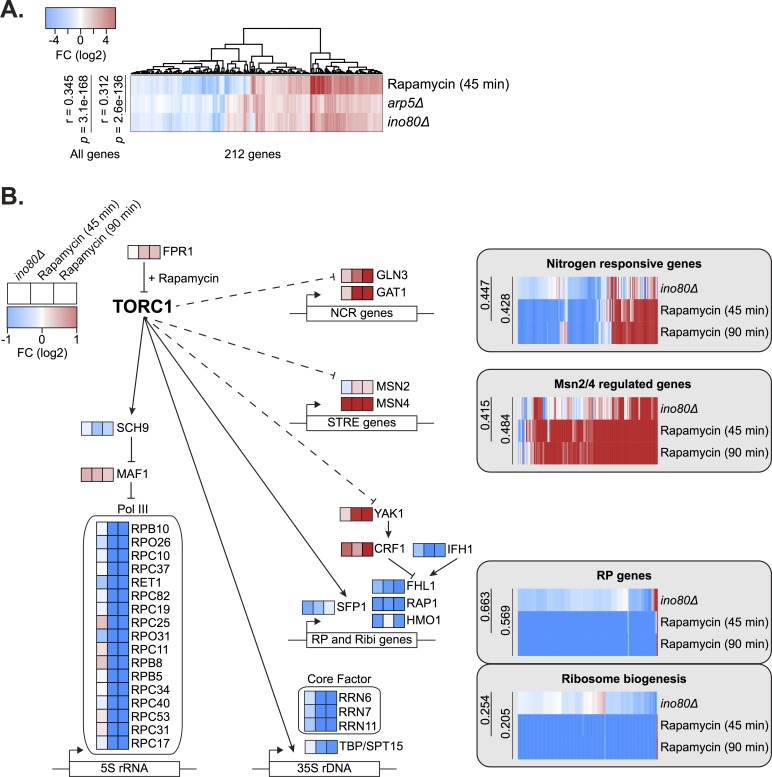
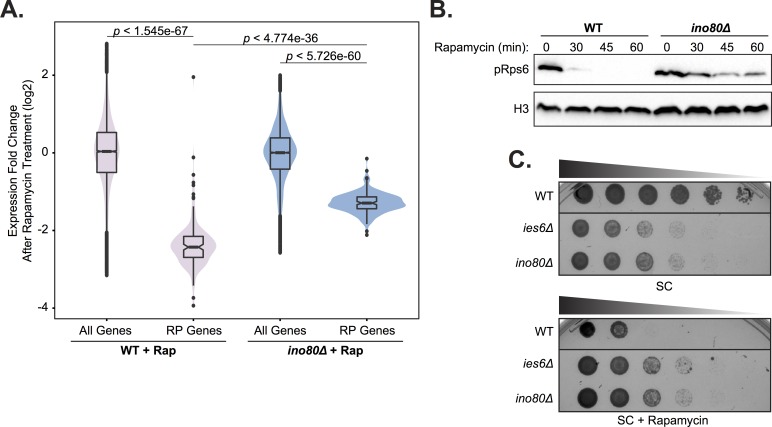
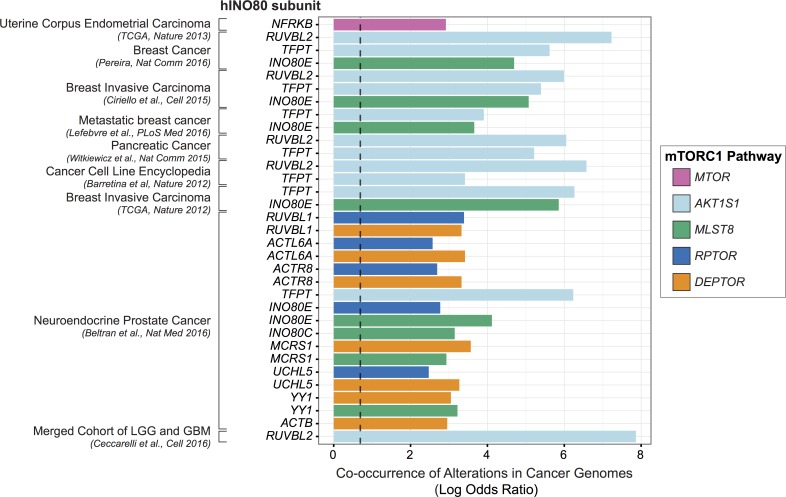
Chromatin remodeling complexes are essential for gene expression programs that coordinate cell function with metabolic status. However, how these remodelers are integrated in metabolic stability pathways is not well known. Here, we report an expansive genetic screen with chromatin remodelers and metabolic regulators in Saccharomyces cerevisiae. We found that, unlike the SWR1 remodeler, the INO80 chromatin remodeling complex is composed of multiple distinct functional subunit modules. We identified a strikingly divergent genetic signature for the Ies6 subunit module that links the INO80 complex to metabolic homeostasis. In particular, mitochondrial maintenance is disrupted in ies6 mutants. INO80 is also needed to communicate TORC1-mediated signaling to chromatin, as ino80 mutants exhibit defective transcriptional profiles and altered histone acetylation of TORC1-responsive genes. Furthermore, comparative analysis reveals subunits of INO80 and mTORC1 have high co-occurrence of alterations in human cancers. Collectively, these results demonstrate that the INO80 complex is a central component of metabolic homeostasis that influences histone acetylation and may contribute to disease when disrupted.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Clapier CR, Cairns BR. The biology of chromatin remodeling complexes. Annu Rev Biochem. 2009;78: 273–304. doi: 10.1146/annurev.biochem.77.062706.153223 - DOI - PubMed
-
- Davis P, Brachmann R. Chromatic Remodeling and Cancer. Cancer Biol Ther. 2003;2: 24–31. Available: http://www.landesbioscience.com/journals/cbt/davis2-1.pdf?origin=publica... - PubMed
-
- de la Serna IL, Ohkawa Y, Imbalzano AN. Chromatin remodelling in mammalian differentiation: lessons from ATP-dependent remodellers. Nat Rev Genet. 2006;7: 461–73. doi: 10.1038/nrg1882 - DOI - PubMed
-
- Wu JI. Diverse functions of ATP-dependent chromatin remodeling complexes in development and cancer. Acta Biochim Biophys Sin (Shanghai). 2012;44: 54–69. doi: 10.1093/abbs/gmr099.Review - DOI - PubMed
-
- Alcid E a., Tsukiyama T. ATP-dependent chromatin remodeling shapes the long noncoding RNA landscape. Genes Dev. 2014;28: 2348–2360. doi: 10.1101/gad.250902.114 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
