

The beta-adrenergic agonist salbutamol modulates neuromuscular junction formation in zebrafish models of human myasthenic syndromes
- PMID: 29462491
- PMCID: PMC5905648
- DOI: 10.1093/hmg/ddy062
The beta-adrenergic agonist salbutamol modulates neuromuscular junction formation in zebrafish models of human myasthenic syndromes
Abstract
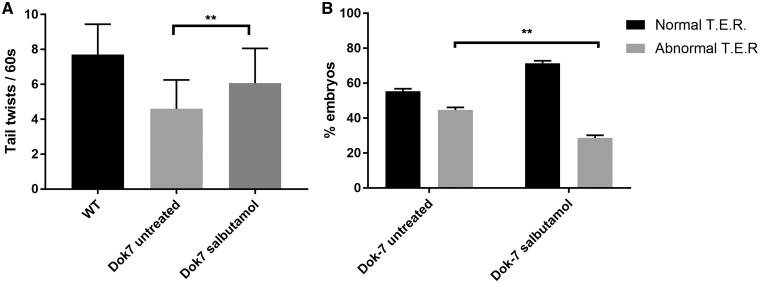
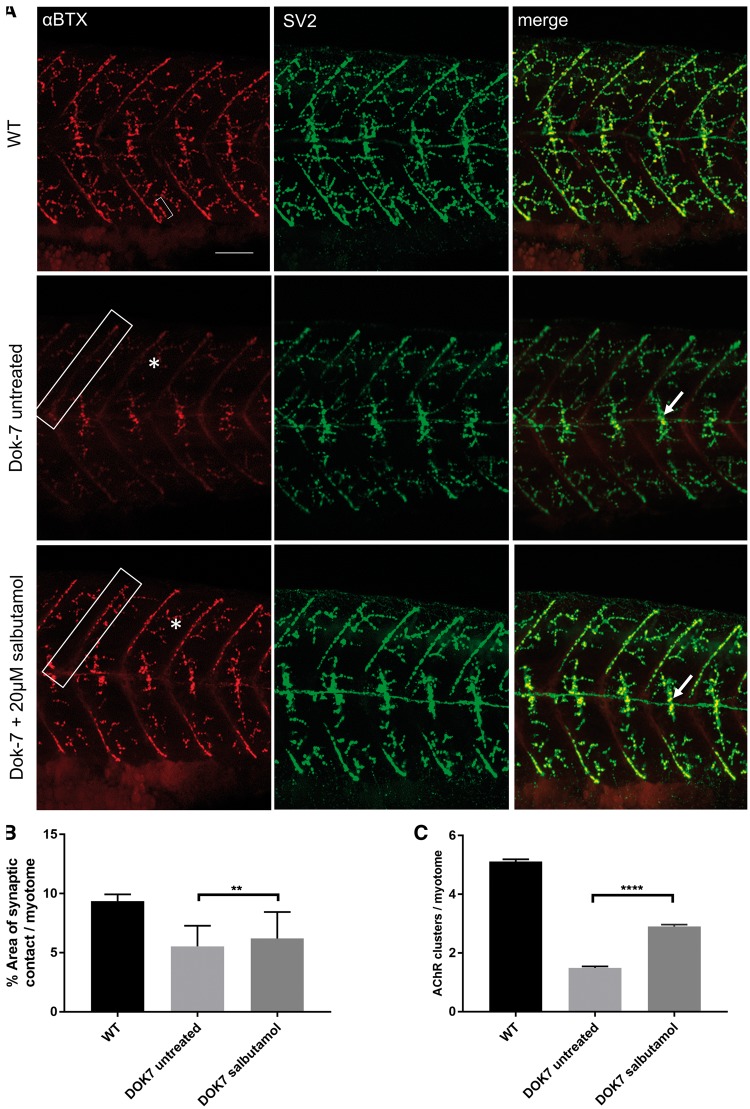
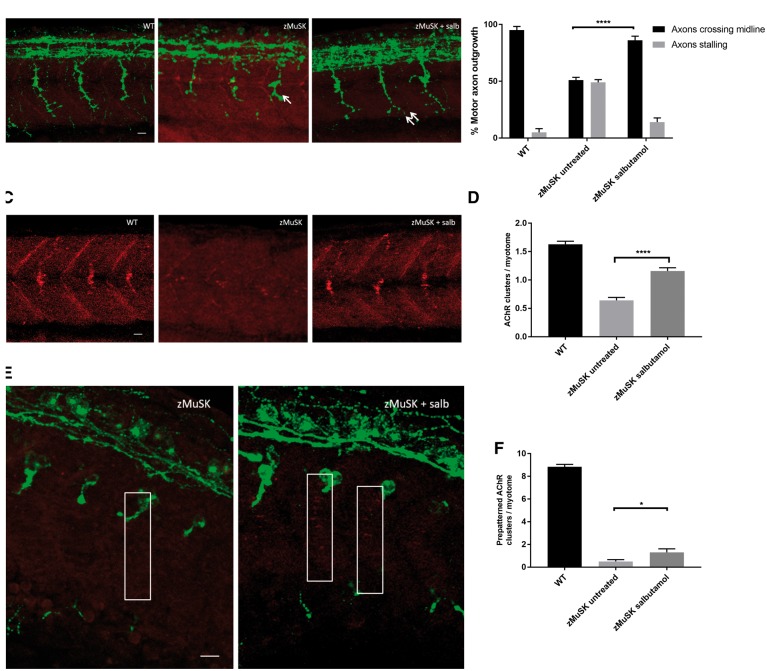
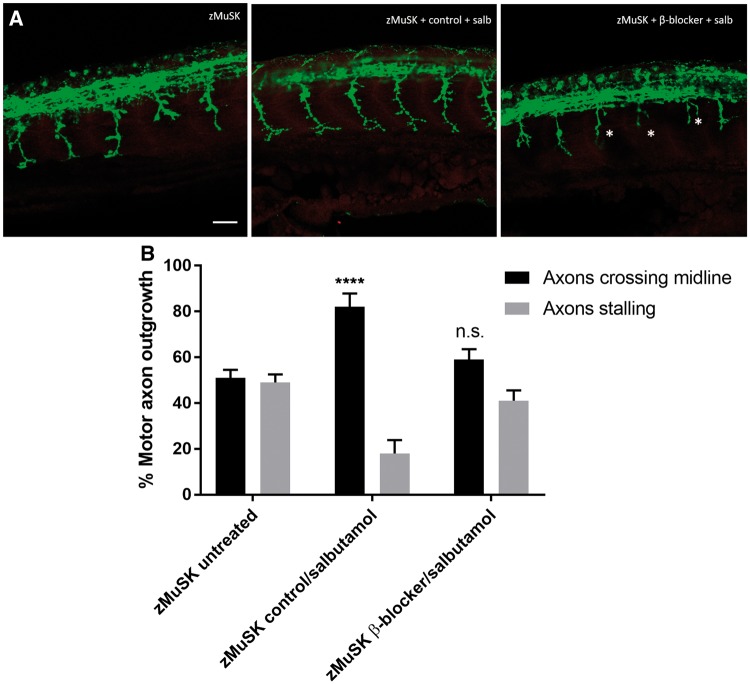
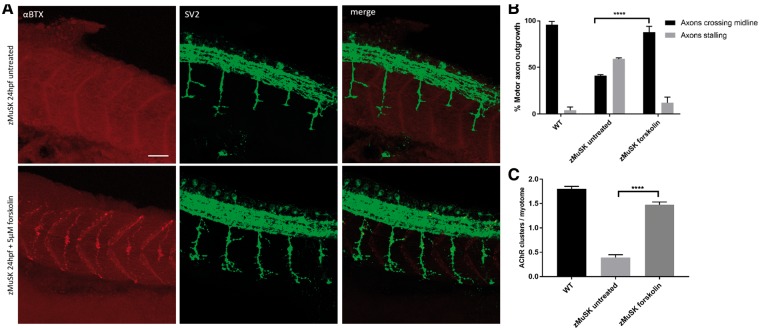
Inherited defects of the neuromuscular junction (NMJ) comprise an increasingly diverse range of disorders, termed congenital myasthenic syndromes (CMS). Therapies acting on the sympathetic nervous system, including the selective β2 adrenergic agonist salbutamol and the α and β adrenergic agonist ephedrine, have become standard treatment for several types of CMS. However, the mechanism of the therapeutic effect of sympathomimetics in these disorders is not understood. Here, we examined the effect of salbutamol on NMJ development using zebrafish with deficiency of the key postsynaptic proteins Dok-7 and MuSK. Treatment with salbutamol reduced motility defects in zebrafish embryos and larvae. In addition, salbutamol lead to morphological improvement of postsynaptic acetycholine receptor (AChR) clustering and size of synaptic contacts in Dok-7-deficient zebrafish. In MuSK-deficient zebrafish, salbutamol treatment reduced motor axon pathfinding defects and partially restored the formation of aneural prepatterned AChRs. In addition, the effects of salbutamol treatment were prevented by pre-treatment with a selective β2 antagonist. Treatment with the cyclic adenosine monophosphate (cAMP) activator forskolin, replicated the effects of salbutamol treatment. These results suggest that sympathomimetics exert a direct effect on neuromuscular synaptogenesis and do so via β2 adrenoceptors and via a cAMP-dependent pathway.
Figures
References
-
- Cohen S.A., Fischbach G.D. (1977) Clusters of acetylcholine receptors located at identified nerve-muscle synapses in vitro. Dev. Biol., 59, 24–38. - PubMed
-
- Kummer T.T., Misgeld T., Sanes J.R. (2006) Assembly of the postsynaptic membrane at the neuromuscular junction: paradigm lost. Curr. Opin. Neurobiol., 16, 74–82. - PubMed
-
- Weatherbee S.D., Anderson K.V., Niswander L.A. (2006) LDL-receptor-related protein 4 is crucial for formation of the neuromuscular junction. Dev. Camb. Engl., 133, 4993–5000. - PubMed
-
- Flanagan-Steet H., Fox M.A., Meyer D., Sanes J.R. (2005) Neuromuscular synapses can form in vivo by incorporation of initially aneural postsynaptic specializations. Development, 132, 4471–4481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
