

Inhibition of autophagy sensitizes cancer cells to Photofrin-based photodynamic therapy
- PMID: 29463237
- PMCID: PMC5819678
- DOI: 10.1186/s12885-018-4126-y
Inhibition of autophagy sensitizes cancer cells to Photofrin-based photodynamic therapy
Abstract
Background: Accumulating evidence suggest that autophagy plays a pivotal role in various anticancer therapies, including photodynamic therapy (PDT), acting as a pro-death or pro-survival mechanism in a context-dependent manner. Therefore, we aimed to determine the role of autophagy in Photofrin-based PDT.
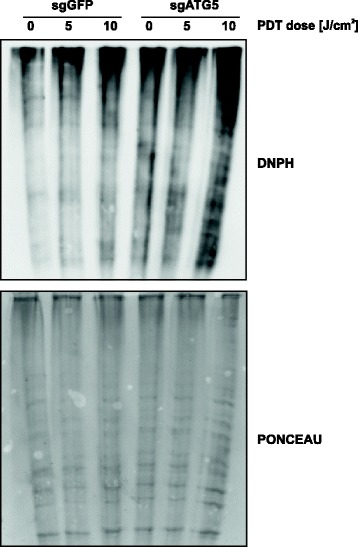
Methods: In vitro cytotoxic/cytostatic effects of PDT were evaluated with crystal violet cell viability assay. Autophagy induction was analyzed by immunoblotting and immunofluorescence using anti-LC3 antibody. Autophagy was inhibited by shRNA-mediated ATG5 knockdown or CRISPR/Cas9-mediated ATG5 knockout. Apoptosis was assessed by flow cytometry analysis of propidium iodide and anexin V-positive cells as well as by detection of cleaved PARP and caspase 3 proteins using immunoblotting. Protein carbonylation was evaluated by the 2,4-dinitrophenylhydrazine (DNPH) method.
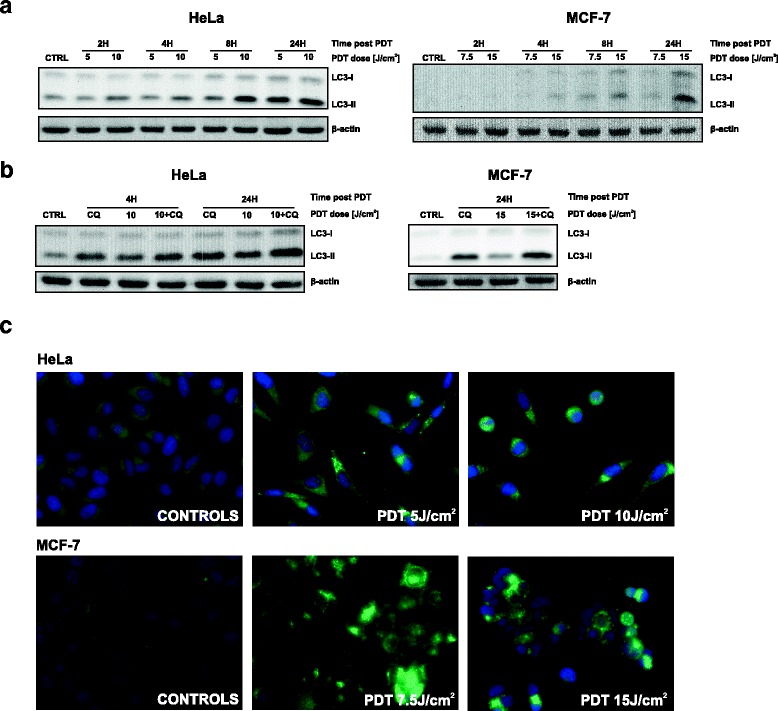
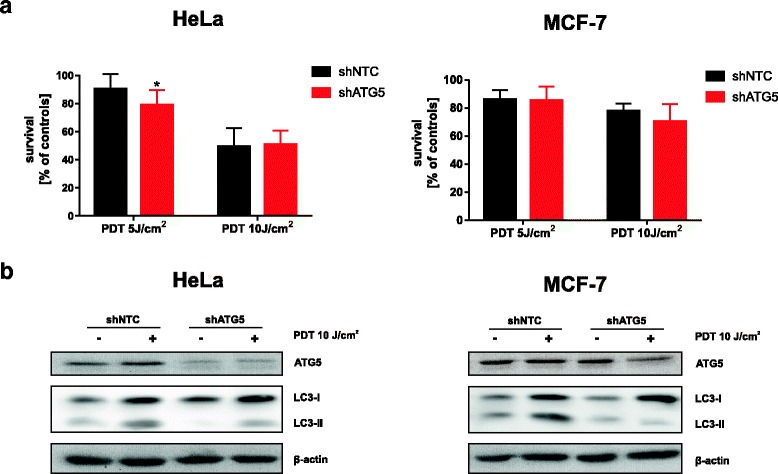
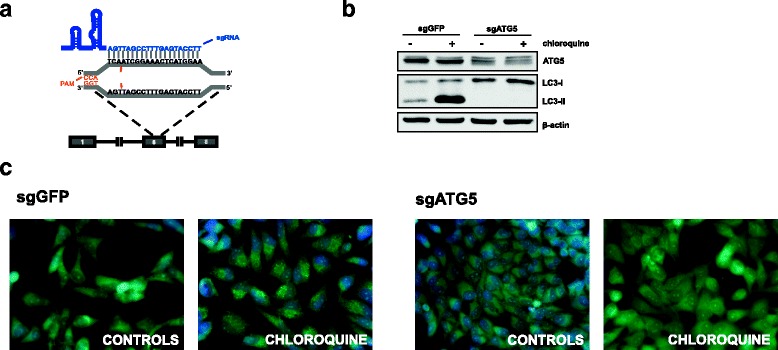
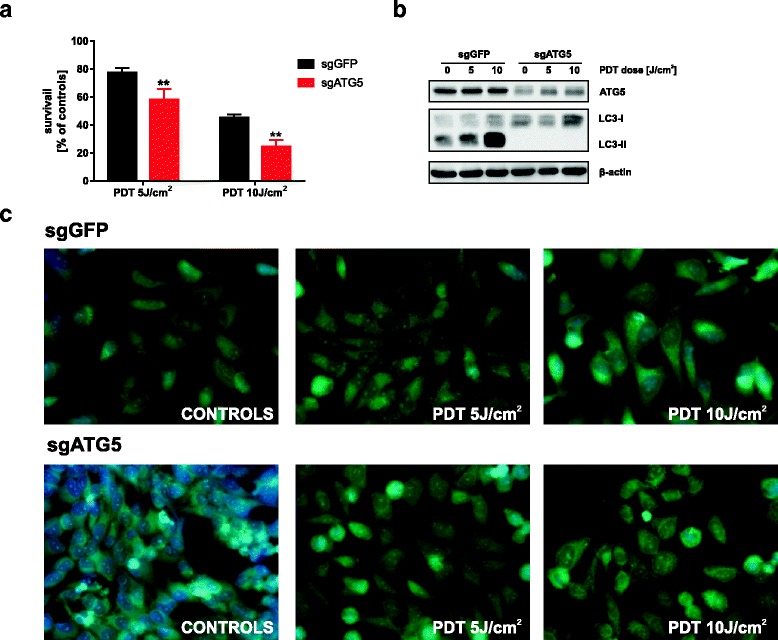
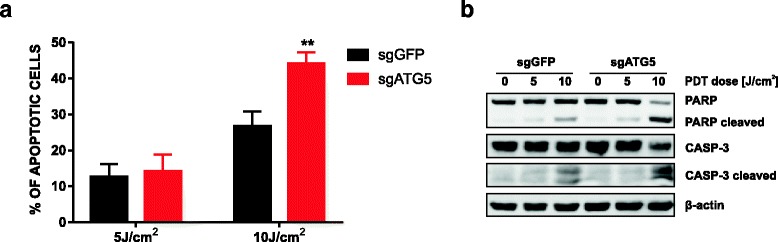
Results: Photofrin-PDT leads to robust autophagy induction in two cancer cell lines, Hela and MCF-7. shRNA-mediated knockdown of ATG5 only partially blocks autophagic response and only marginally affects the sensitivity of Hela and MCF-7 cells to PDT. ATG5 knockout in HeLa cell line utilizing CRISPR/Cas9 genome editing results in increased PDT-mediated cytotoxicity, which is accompanied by an enhanced apoptotic response and increased accumulation of carbonylated proteins.
Conclusions: Altogether, these observations imply that autophagy contributes to Photofrin-PDT resistance by enabling clearance of carbonylated and other damaged proteins. Therefore, autophagy inhibition may serve as a strategy to improve PDT efficacy.
Keywords: ATG5; Autophagy; CRISR/Cas-9; Photodynamic therapy; Photofrin.
Conflict of interest statement
Ethics approval and consent to participate
There was no research involving human or animal subjects in the study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interest.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
