

Transcriptional factor ICER promotes glutaminolysis and the generation of Th17 cells
- PMID: 29463741
- PMCID: PMC5877961
- DOI: 10.1073/pnas.1714717115
Transcriptional factor ICER promotes glutaminolysis and the generation of Th17 cells
Abstract
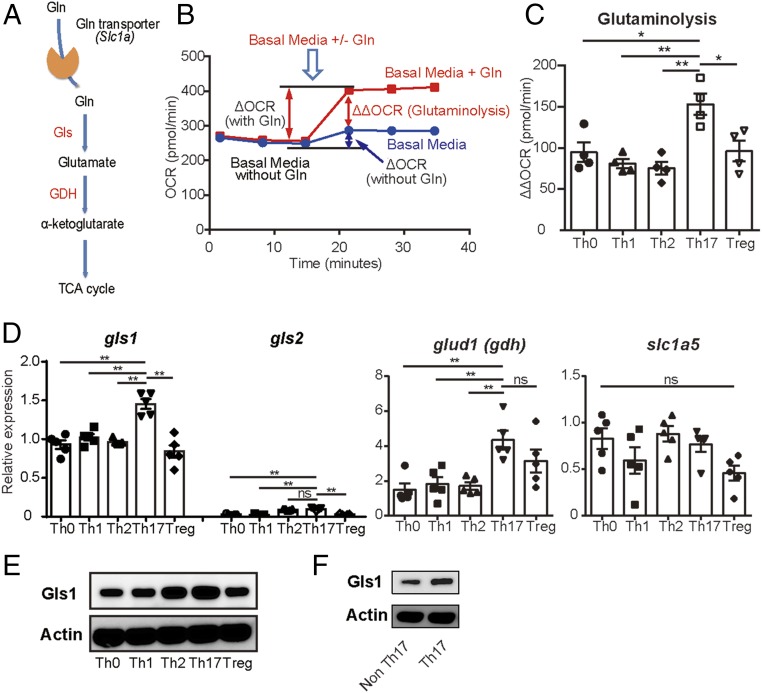
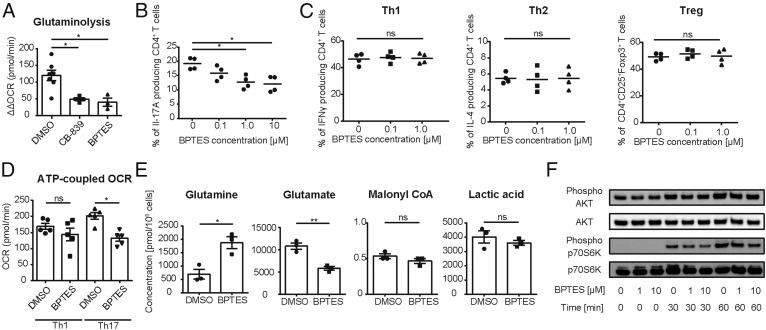
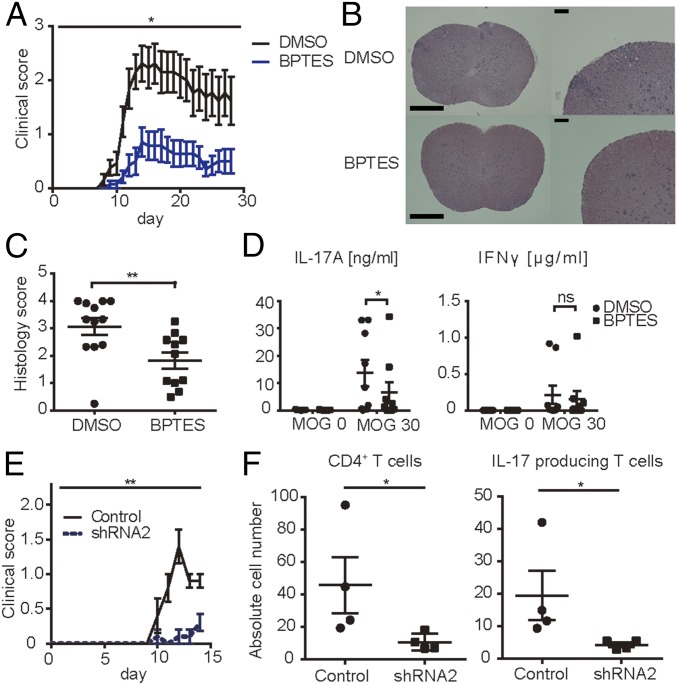
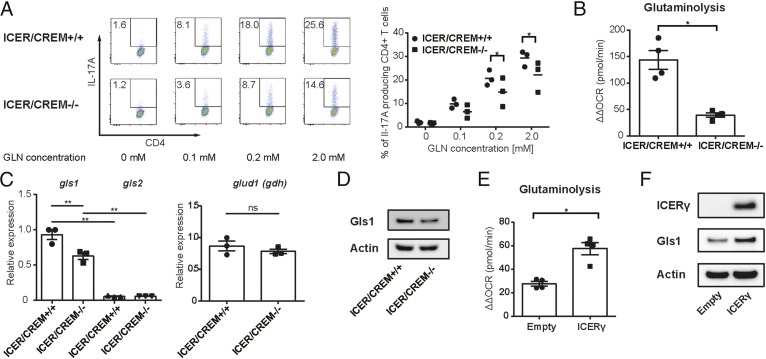
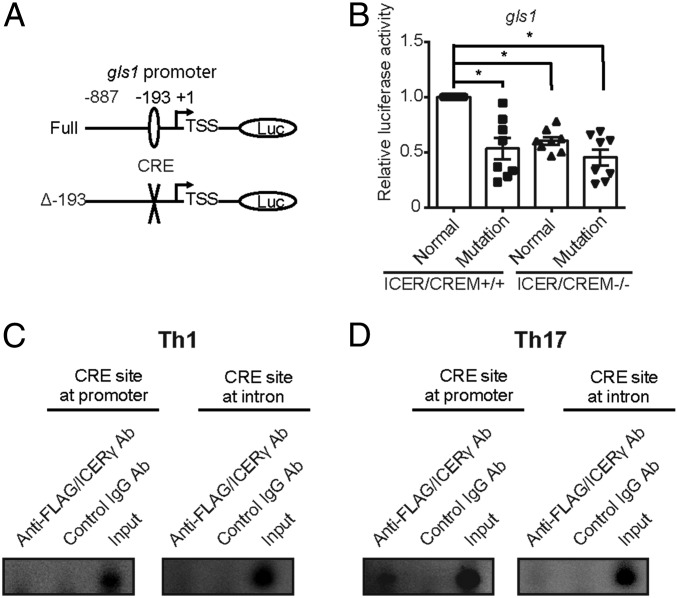
Glutaminolysis is a well-known source of energy for effector T cells but its contribution to each T cell subset and the mechanisms which are responsible for the control of involved metabolic enzymes are not fully understood. We report that Th17 but not Th1, Th2, or Treg cell induction in vitro depends on glutaminolysis and the up-regulation of glutaminase 1 (Gls1), the first enzyme in the glutaminolysis pathway. Both pharmacological and siRNA-based selective inhibition of Gls1 reduced in vitro Th17 differentiation and reduced the CD3/TCR-mediated increase of the mammalian target of rapamycin complex 1 activity. Treatment of mice with a Gls1 inhibitor ameliorated experimental autoimmune encephalomyelitis. Furthermore, RAG1-deficient mice that received Gls1-shRNA-transfected 2D2 T cells had reduced experimental autoimmune encephalomyelitis scores compared with those that received control-shRNA-treated cells. Next we found that T cells deficient in inducible cAMP early repressor (ICER), a transcriptional factor known to promote Th17 differentiation, display reduced activity of oxidative phosphorylation rates in the presence of glutamine and reduced Gls1 expression, both of which could be restored by ICER overexpression. Finally, we demonstrate that ICER binds to the gls1 promoter directly and increases its activity. These findings demonstrate the importance of glutaminolysis in the generation of Th17 and the direct control of Gls1 activity by the IL-17-promoting transcription factor ICER. Pharmaceutical modulation of the glutaminolysis pathway should be considered to control Th17-mediated pathology.
Keywords: ICER; Th17; autoimmunity; glutaminase 1; glutaminolysis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 cells. Annu Rev Immunol. 2009;27:485–517. - PubMed
-
- Di Cesare A, Di Meglio P, Nestle FO. The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J Invest Dermatol. 2009;129:1339–1350. - PubMed
-
- Ooi JD, Kitching AR, Holdsworth SR. Review: T helper 17 cells: Their role in glomerulonephritis. Nephrology (Carlton) 2010;15:513–521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
