

Danger Changes the Way the Mammalian Brain Stores Information About Innocuous Events: A Study of Sensory Preconditioning in Rats
- PMID: 29464195
- PMCID: PMC5815846
- DOI: 10.1523/ENEURO.0381-17.2017
Danger Changes the Way the Mammalian Brain Stores Information About Innocuous Events: A Study of Sensory Preconditioning in Rats
Abstract
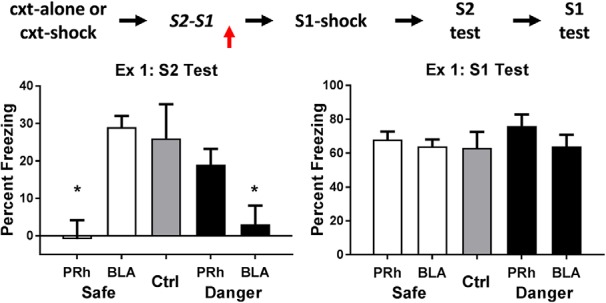
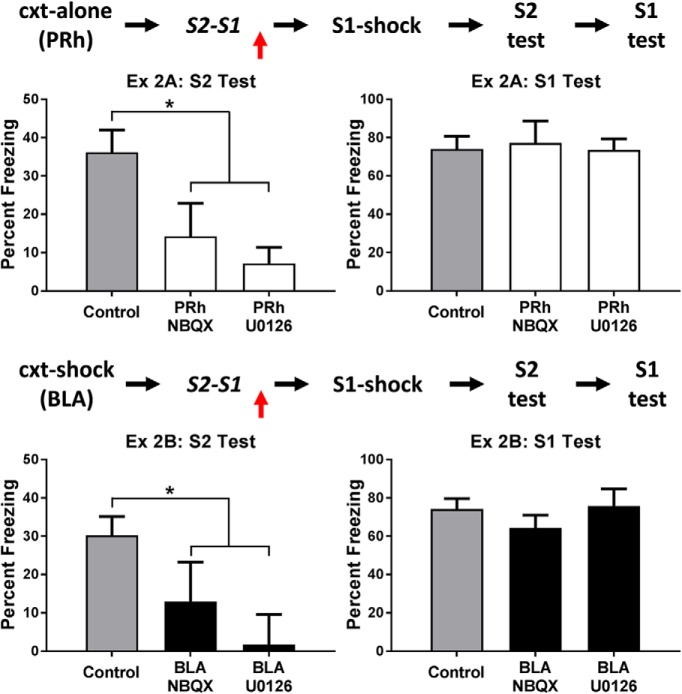
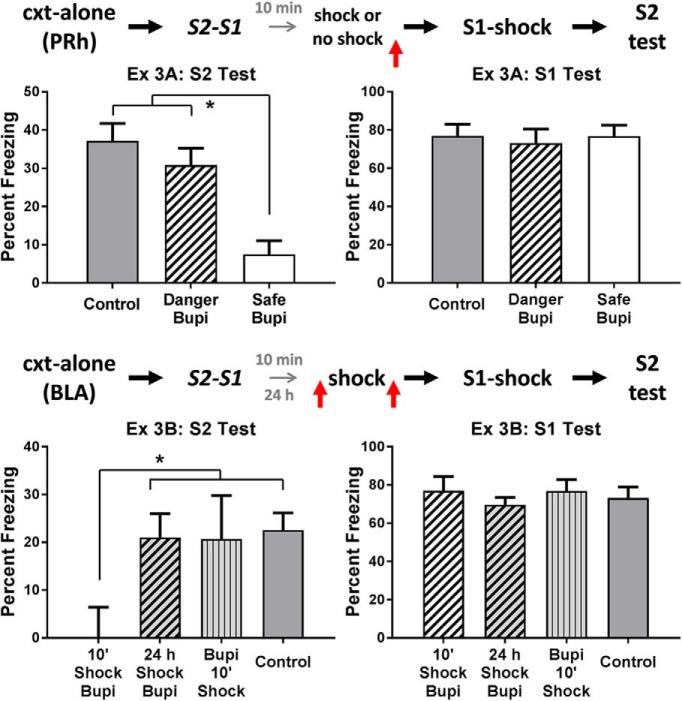
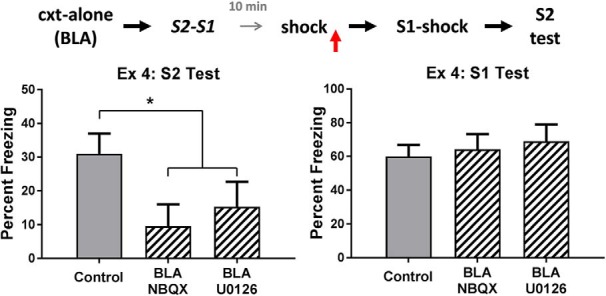
The amygdala is a critical substrate for learning about cues that signal danger. Less is known about its role in processing innocuous or background information. The present study addressed this question using a sensory preconditioning protocol in male rats. In each experiment, rats were exposed to pairings of two innocuous stimuli in stage 1, S2 and S1, and then to pairings of S1 and shock in stage 2. As a consequence of this training, control rats displayed defensive reactions (freezing) when tested with both S2 and S1. The freezing to S2 is a product of two associations formed in training: an S2-S1 association in stage 1 and an S1-shock association in stage 2. We examined the roles of two medial temporal lobe (MTL) structures in consolidation of the S2-S1 association: the perirhinal cortex (PRh) and basolateral complex of the amygdala (BLA). When the S2-S1 association formed in a safe context, its consolidation required neuronal activity in the PRh (but not BLA), including activation of AMPA receptors and MAPK signaling. In contrast, when the S2-S1 association formed in a dangerous context, or when the context was rendered dangerous immediately after the association had formed, its consolidation required neuronal activity in the BLA (but not PRh), including activation of AMPA receptors and MAPK signaling. These roles of the PRh and BLA show that danger changes the way the mammalian brain stores information about innocuous events. They are discussed with respect to danger-induced changes in stimulus processing.
Keywords: amygdala; associative learning; consolidation; danger; memory; sensory preconditioning.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials