

Subpopulations of vomeronasal sensory neurons with coordinated coexpression of type 2 vomeronasal receptor genes are differentially dependent on Vmn2r1
- PMID: 29465786
- PMCID: PMC5947554
- DOI: 10.1111/ejn.13875
Subpopulations of vomeronasal sensory neurons with coordinated coexpression of type 2 vomeronasal receptor genes are differentially dependent on Vmn2r1
Abstract
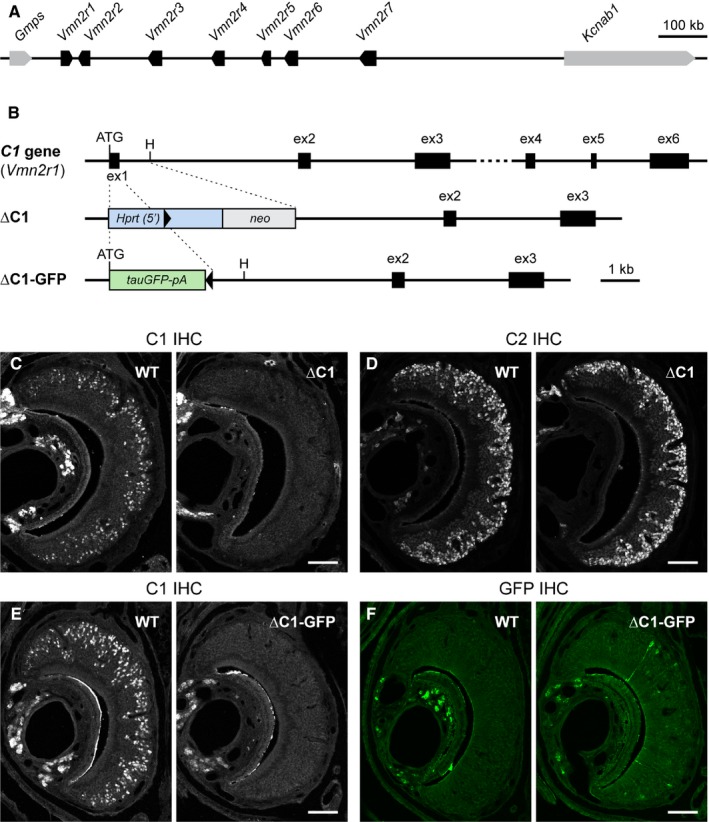
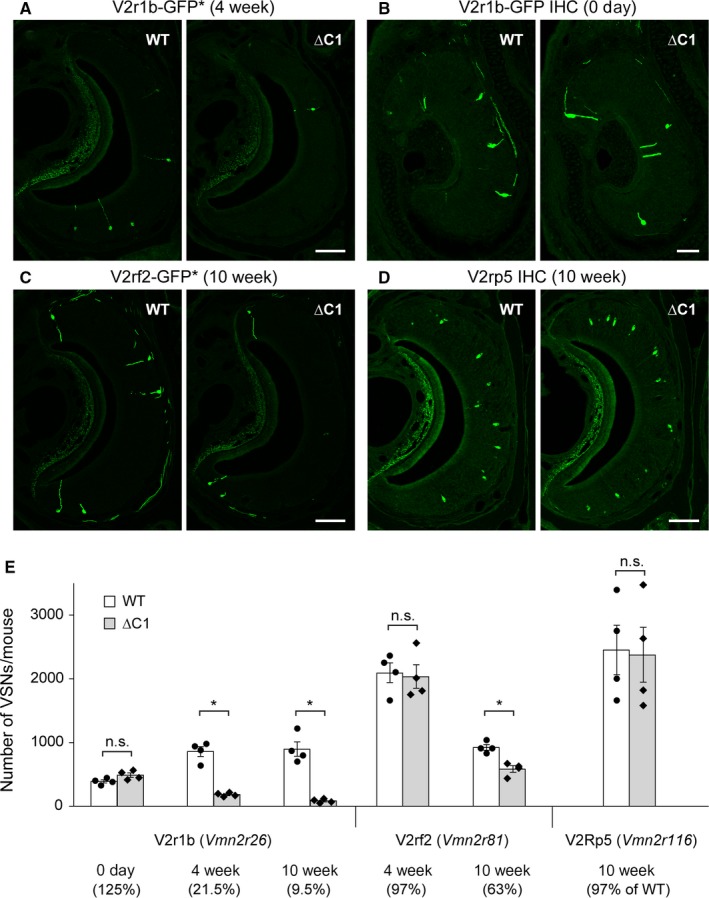
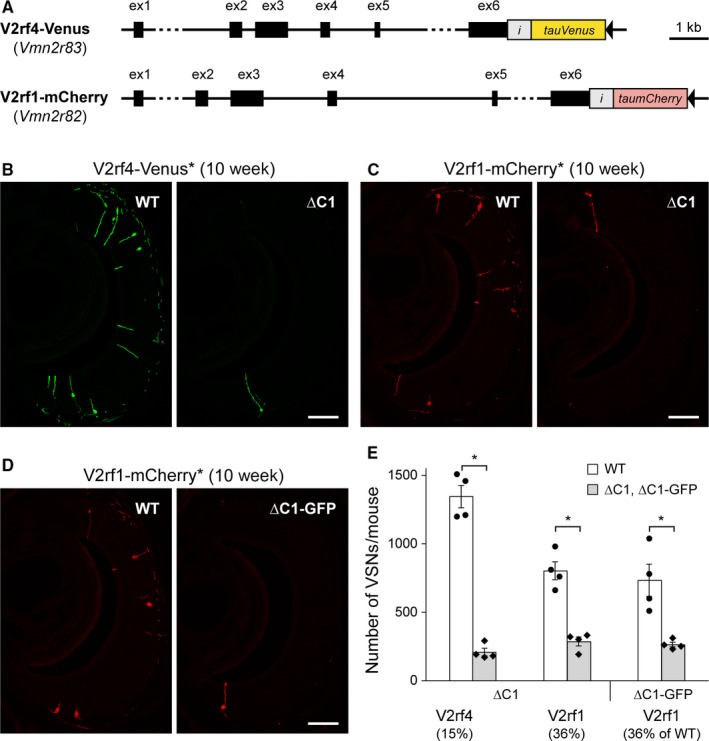
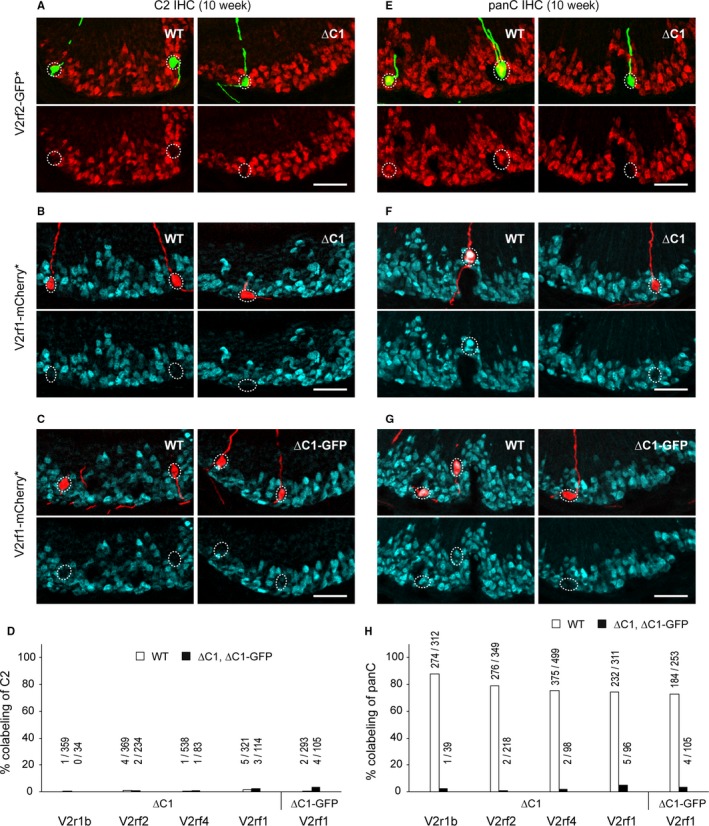
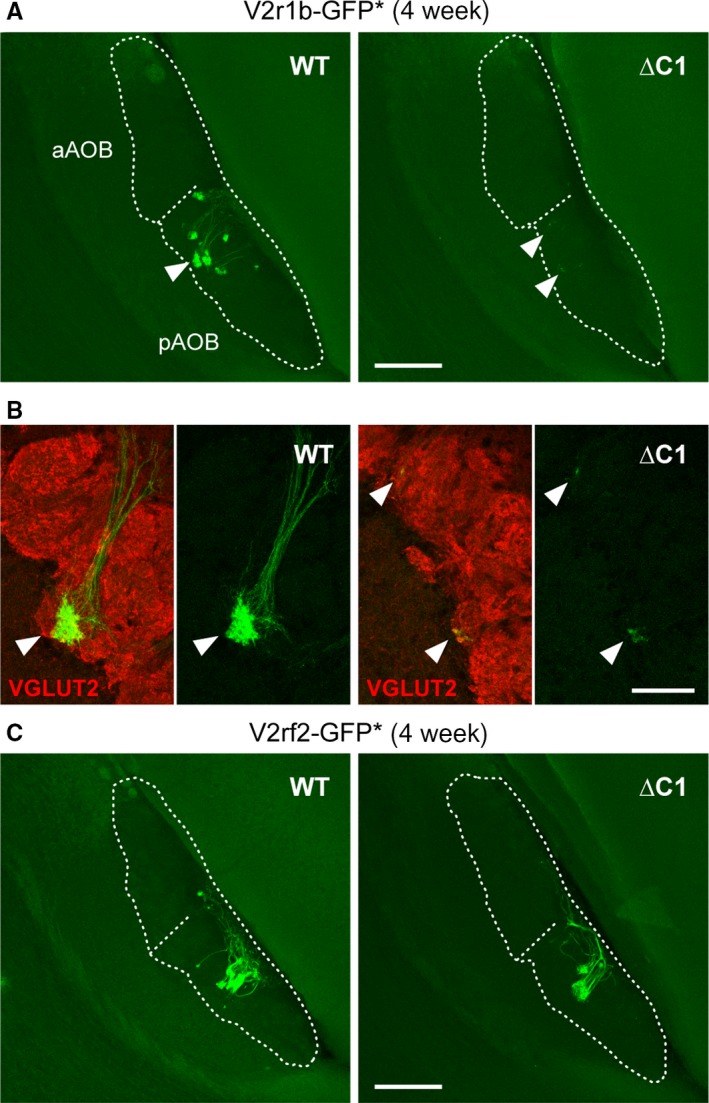
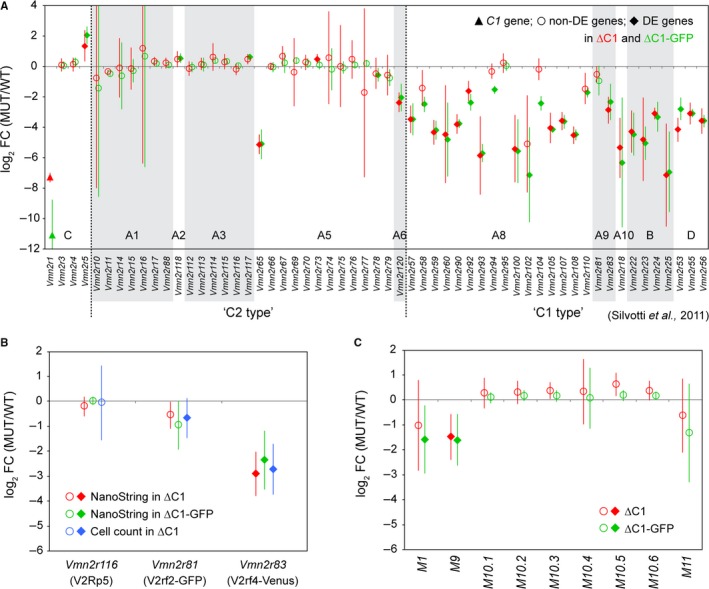
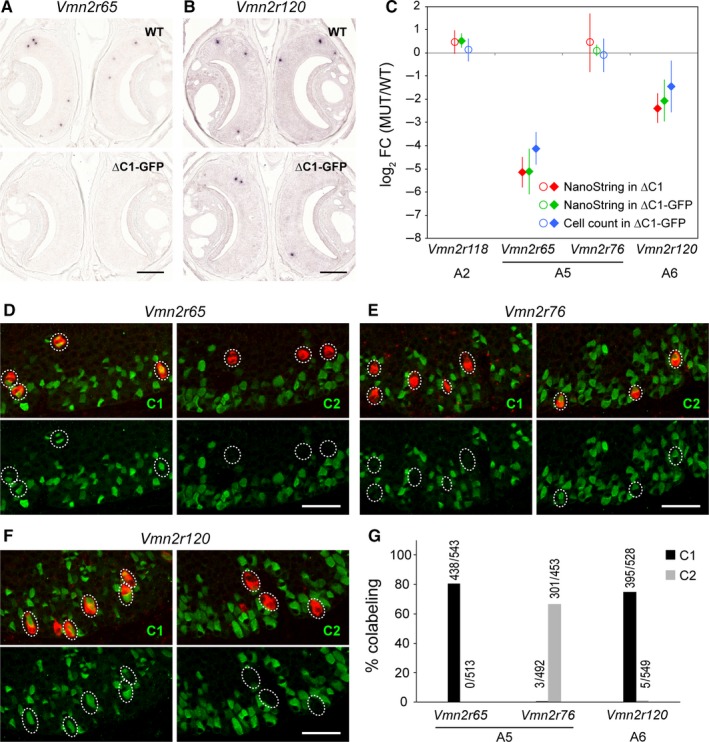
The mouse vomeronasal organ is specialized in the detection of pheromones. Vomeronasal sensory neurons (VSNs) express chemosensory receptors of two large gene repertoires, V1R and V2R, which encode G-protein-coupled receptors. Phylogenetically, four families of V2R genes can be discerned as follows: A, B, C, and D. VSNs located in the basal layer of the vomeronasal epithelium coordinately coexpress V2R genes from two families: Approximately half of basal VSNs coexpress Vmn2r1 of family C with a single V2R gene of family A8-10, B, or D ('C1 type of V2Rs'), and the other half coexpress Vmn2r2 through Vmn2r7 of family C with a single V2R gene of family A1-6 ('C2 type V2Rs'). The regulatory mechanisms of the coordinated coexpression of V2Rs from two families remain poorly understood. Here, we have generated two mouse strains carrying a knockout mutation in Vmn2r1 by gene targeting in embryonic stem cells. These mutations cause a differential decrease in the numbers of VSNs expressing a given C1 type of V2R. There is no compensatory expression of Vmn2r2 through Vmn2r7. VSN axons coalesce into glomeruli in the appropriate region of the accessory olfactory bulb in the absence of Vmn2r1. Gene expression profiling by NanoString reveals a differential and graded decrease in the expression levels across C1 type of V2Rs. There is no change in the expression levels of C2 type of V2Rs, with two exceptions that we reclassified as C1 type. Thus, there appears to be a fixed probability of gene choice for a given C2 type of V2R.
Keywords: V2R; accessory olfactory system; gene expression; gene regulation; pheromone.
© 2018 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures
References
-
- Belluscio, L. , Koentges, G. , Axel, R. & Dulac, C. (1999) A map of pheromone receptor activation in the mammalian brain. Cell, 97, 209–220. - PubMed
-
- Benjamini, Y. & Yekutieli, D. (2001) The control of the false discovery rate in multiple testing under dependency. Ann. Stat., 29, 1165–1188.
-
- Chamero, P. , Marton, T.F. , Logan, D.W. , Flanagan, K. , Cruz, J.R. , Saghatelian, A. , Cravatt, B.F. & Stowers, L. (2007) Identification of protein pheromones that promote aggressive behaviour. Nature, 450, 899–902. - PubMed
-
- Chamero, P. , Leinders‐Zufall, T. & Zufall, F. (2012) From genes to social communication: molecular sensing by the vomeronasal organ. Trends Neurosci., 35, 597–606. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
