

Bioerosion by pit-forming, temperate-reef sea urchins: History, rates and broader implications
- PMID: 29466357
- PMCID: PMC5821313
- DOI: 10.1371/journal.pone.0191278
Bioerosion by pit-forming, temperate-reef sea urchins: History, rates and broader implications
Abstract
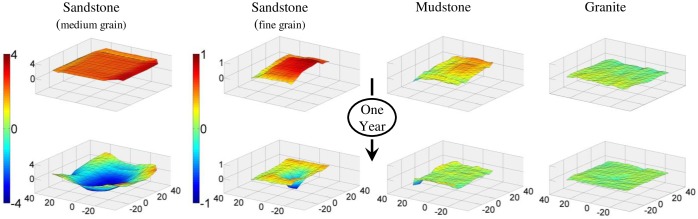
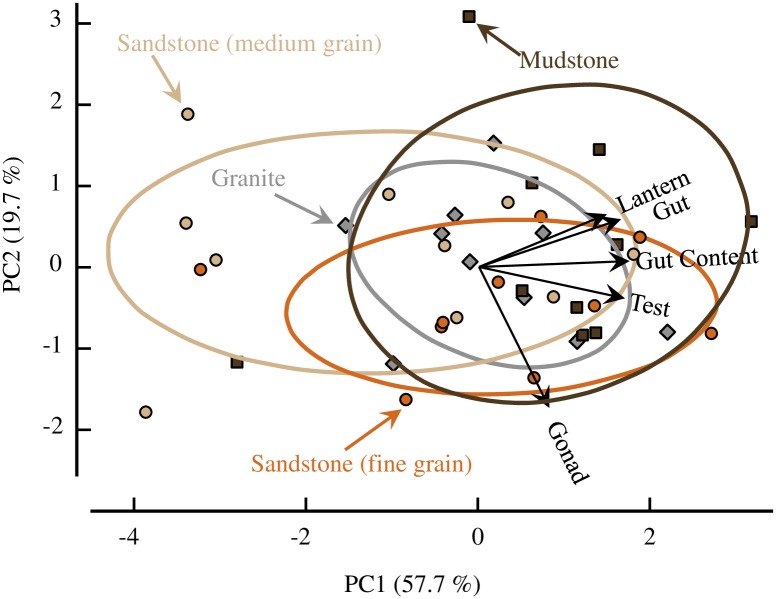
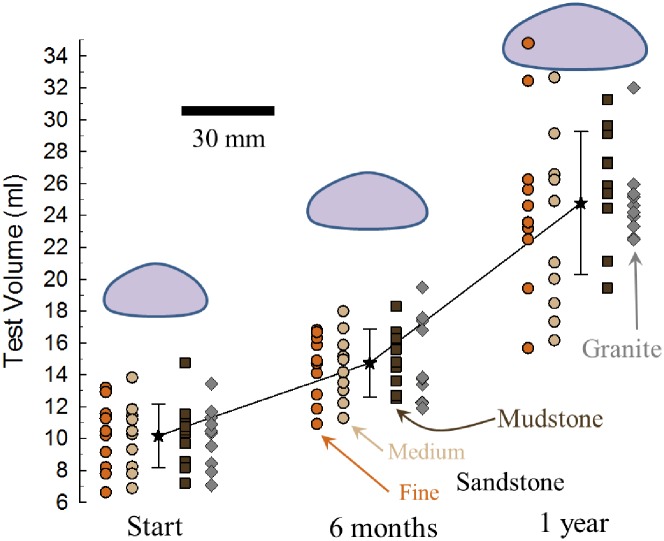
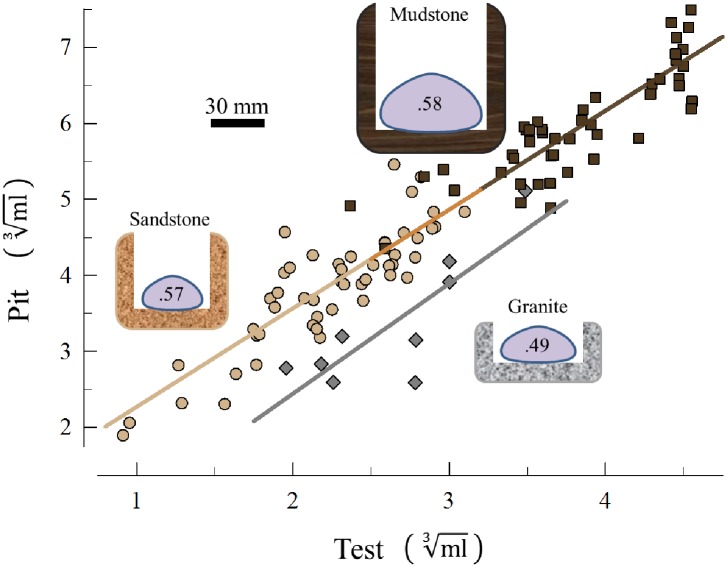
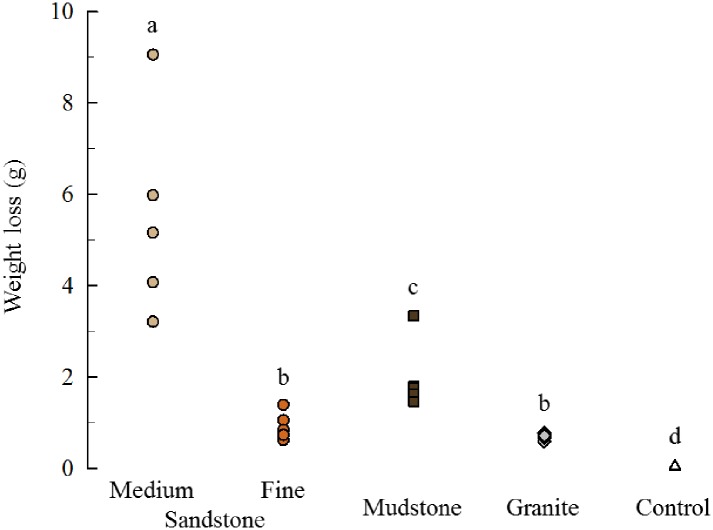
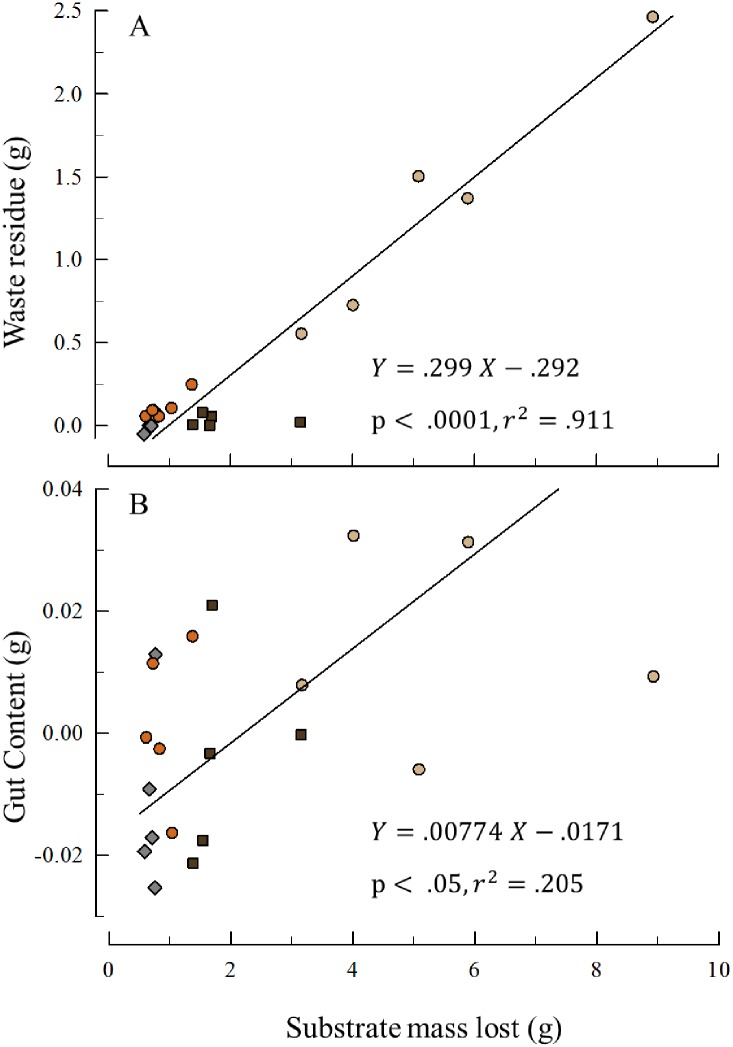
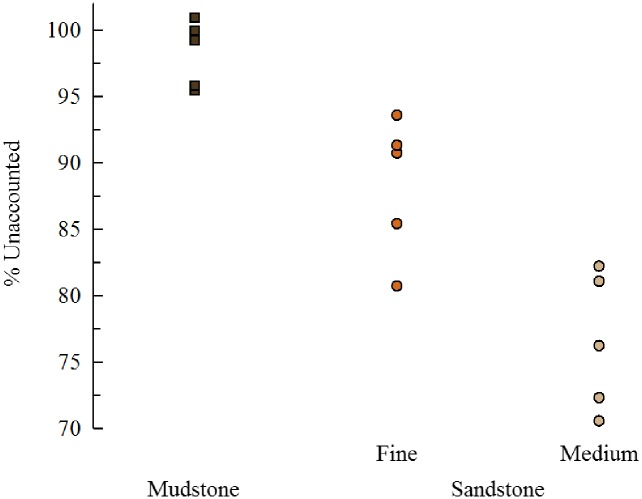
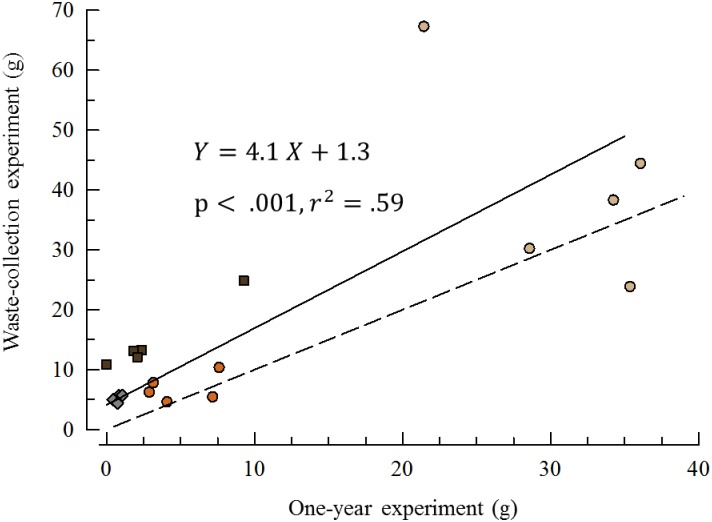
Sea urchins are dominant members of rocky temperate reefs around the world. They often occur in cavities within the rock, and fit so tightly, it is natural to assume they sculpted these "pits." However, there are no experimental data demonstrating they bore pits. If they do, what are the rates and consequences of bioerosion to nearshore systems? We sampled purple sea urchins, Strongylocentrotus purpuratus, from sites with four rock types, three sedimentary (two sandstones and one mudstone) and one metamorphic (granite). A year-long experiment showed urchins excavated depressions on sedimentary rocks in just months. The rate of pit formation varied with rock type and ranged from <5 yr for medium-grain sandstone to >100 yr for granite. In the field, there were differences in pit size and shapes of the urchins (height:diameter ratio). The pits were shallow and urchins flatter at the granite site, and the pits were deeper and urchins taller at the sedimentary sites. Although overall pit sizes were larger on mudstone than on sandstone, urchin size accounted for this difference. A second, short-term experiment, showed the primary mechanism for bioerosion was ingestion of the substratum. This experiment eliminated potential confounding factors of the year-long experiment and yielded higher bioerosion rates. Given the high densities of urchins, large amounts of rock can be converted to sediment over short time periods. Urchins on sandstone can excavate as much as 11.4 kg m-2 yr-1. On a broader geographic scale, sediment production can exceed 100 t ha-1 yr-1, and across their range, their combined bioerosion is comparable to the sediment load of many rivers. The phase shift between urchin barrens and kelp bed habitats in the North Pacific is controlled by the trophic cascade of sea otters. By limiting urchin populations, these apex predators also may indirectly control a substantial component of coastal rates of bioerosion.
Conflict of interest statement
Figures


References
-
- Harrold C, Pearse JS. The ecological role of echinoderms in kelp forests In: Jangoux M, Lawrence JM, editors. Echioderm Studies 2. Rotterdam: A. A. Balkema; 1987. pp. 137–234.
-
- Himmelman JH, Cardinal A, Bourget E. Community development following removal of urchins, Strongylocentrotus droebachiensis, from the rocky subtidal zone of the St. Lawrence Estuary, Eastern Canada. Oecologia. 1983;59:27–39. doi: 10.1007/BF00388068 - DOI - PubMed
-
- Lawrence JM. On the relationships between marine plants and sea urchins. Oceanogr Mar Biol Annu Rev. 1975;13:213–86.
-
- Paine RT. Food webs: linkage, interaction strength and community infrastructure. J Anim Ecol. 1980;49(3):667–85.
-
- Steneck RS. Sea urchins as drivers of shallow benthic marine community structure. In: Lawrence JM, editor. Sea Urchins: Biology and Ecology. 2013;38:195–212.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
