

T Cells in Nonlymphoid Tissues Give Rise to Lymph-Node-Resident Memory T Cells
- PMID: 29466758
- PMCID: PMC5828517
- DOI: 10.1016/j.immuni.2018.01.015
T Cells in Nonlymphoid Tissues Give Rise to Lymph-Node-Resident Memory T Cells
Abstract
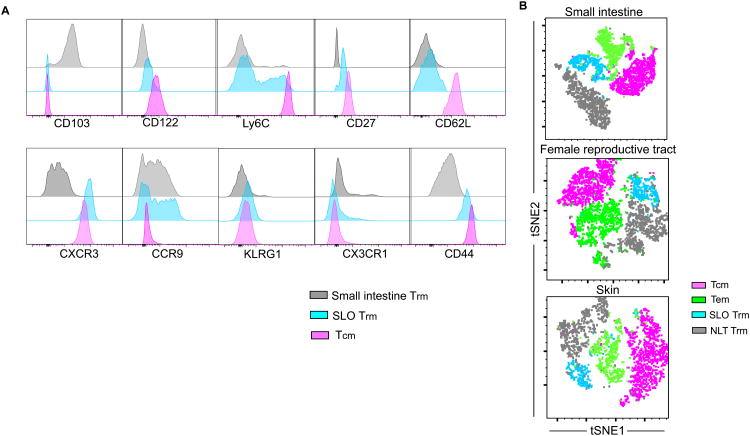
Immunosurveillance of secondary lymphoid organs (SLO) is performed by central memory T cells that recirculate through blood. Resident memory T (Trm) cells remain parked in nonlymphoid tissues and often stably express CD69. We recently identified Trm cells within SLO, but the origin and phenotype of these cells remains unclear. Using parabiosis of "dirty" mice, we found that CD69 expression is insufficient to infer stable residence of SLO Trm cells. Restimulation of nonlymphoid memory CD8+ T cells within the skin or mucosa resulted in a substantial increase in bona fide Trm cells specifically within draining lymph nodes. SLO Trm cells derived from emigrants from nonlymphoid tissues and shared some transcriptional and phenotypic signatures associated with nonlymphoid Trm cells. These data indicate that nonlymphoid cells can give rise to SLO Trm cells and suggest vaccination strategies by which memory CD8+ T cell immunosurveillance can be regionalized to specific lymph nodes.
Keywords: local recall immunization; non-lymphoid tissue; regionalized immunosurveillance; resident memory T cells; secondary lymphoid organs.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures






References
-
- Beura LK, Hamilton SE, Bi K, Schenkel JM, Odumade OA, Casey KA, Thompson EA, Fraser KA, Rosato PC, Filali-Mouhim A, Sekaly RP, Jenkins MK, Vezys V, Haining WN, Jameson SC, Masopust D. Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature. 2016;532:512–6. - PMC - PubMed
-
- Beura LK, Mitchell JS, Thompson EA, Schenkel JM, Mohammed J, Wijeyesinghe S, Fonseca R, Burbach BJ, Hickman HD, Vezys V, Fife BT, Masopust D. Intravital mucosal imaging of CD8+ resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory. Nat Immunol. 2018;19:173–182. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
