

apterous A specifies dorsal wing patterns and sexual traits in butterflies
- PMID: 29467265
- PMCID: PMC5832707
- DOI: 10.1098/rspb.2017.2685
apterous A specifies dorsal wing patterns and sexual traits in butterflies
Abstract
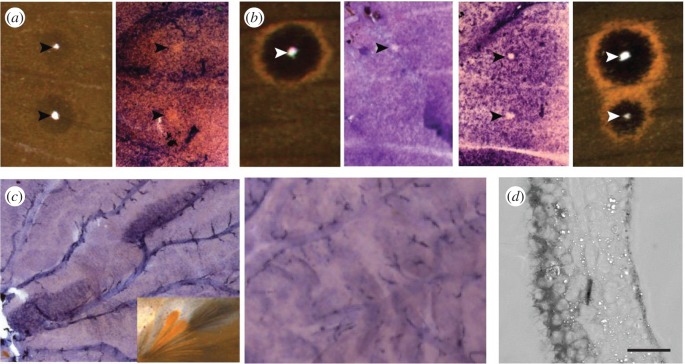
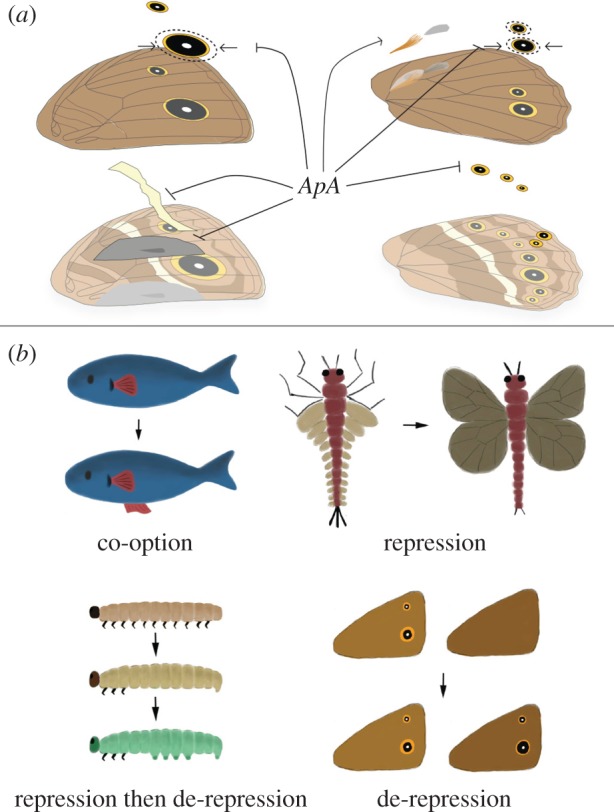
Butterflies have evolved different colour patterns on their dorsal and ventral wing surfaces to serve different signalling functions, yet the developmental mechanisms controlling surface-specific patterning are still unknown. Here, we mutate both copies of the transcription factor apterous in Bicyclus anynana butterflies using CRISPR/Cas9 and show that apterous A, expressed dorsally, functions both as a repressor and modifier of ventral wing colour patterns, as well as a promoter of dorsal sexual ornaments in males. We propose that the surface-specific diversification of wing patterns in butterflies proceeded via the co-option of apterous A or its downstream effectors into various gene regulatory networks involved in the differentiation of discrete wing traits. Further, interactions between apterous and sex-specific factors such as doublesex may have contributed to the origin of sexually dimorphic surface-specific patterns. Finally, we discuss the evolution of eyespot number diversity in the family Nymphalidae within the context of developmental constraints due to apterous regulation.
Keywords: apterous; butterfly wing patterns; developmental constraints; dorsal–ventral differentiation; eyespot repression.
© 2018 The Authors.
Conflict of interest statement
We declare we have no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials