

GRAM domain proteins specialize functionally distinct ER-PM contact sites in human cells
- PMID: 29469807
- PMCID: PMC5823543
- DOI: 10.7554/eLife.31019
GRAM domain proteins specialize functionally distinct ER-PM contact sites in human cells
Abstract
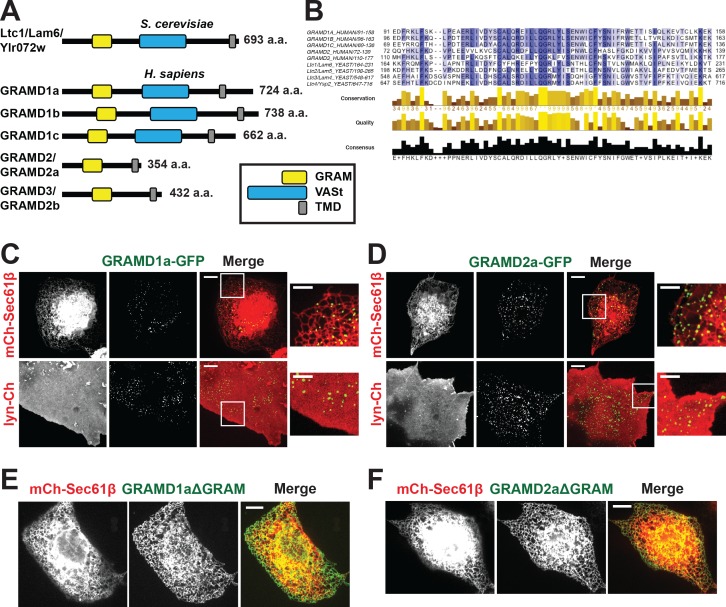
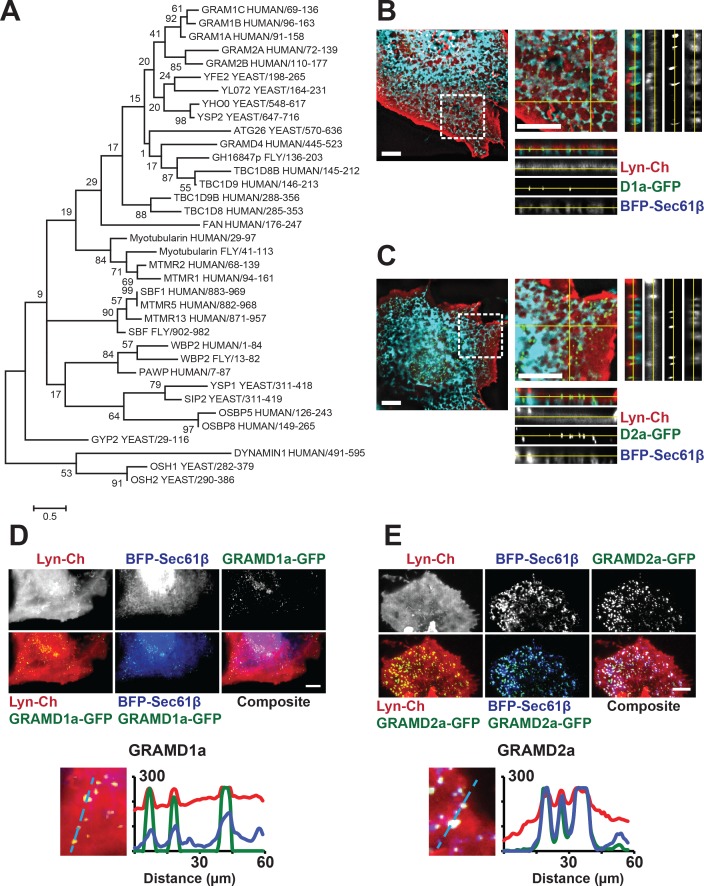
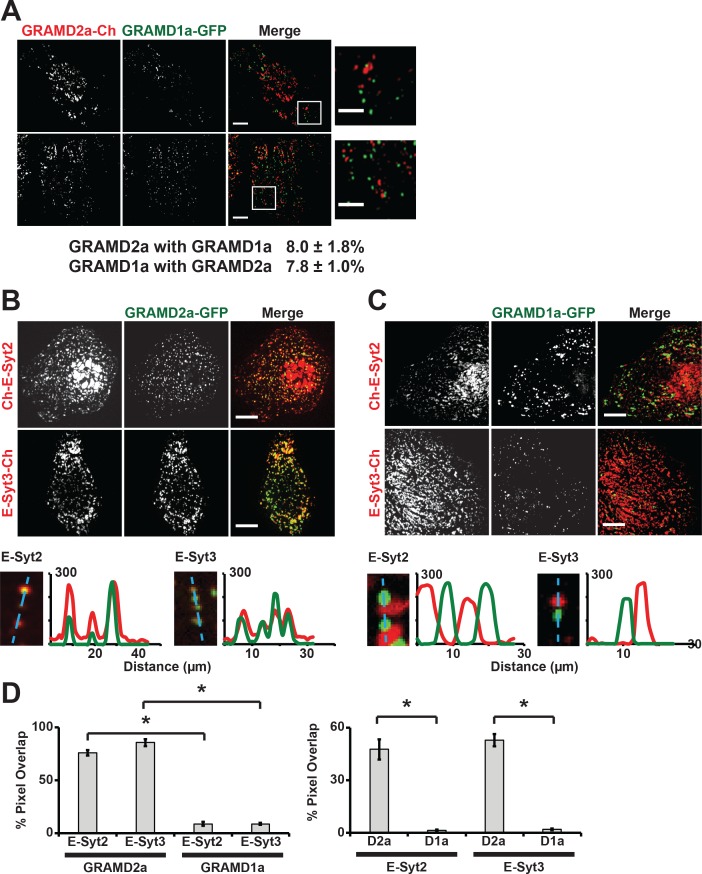
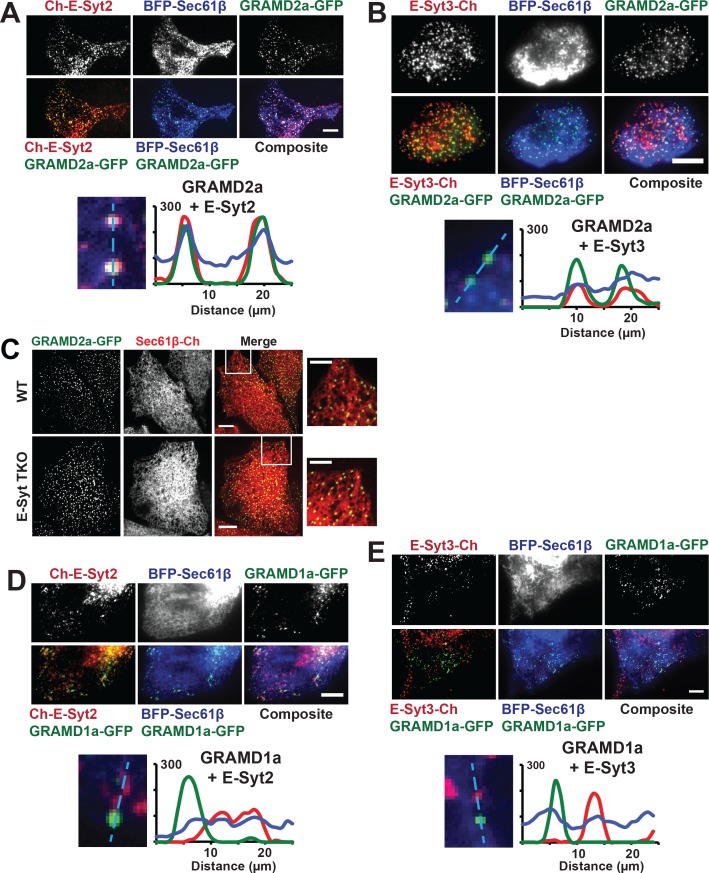
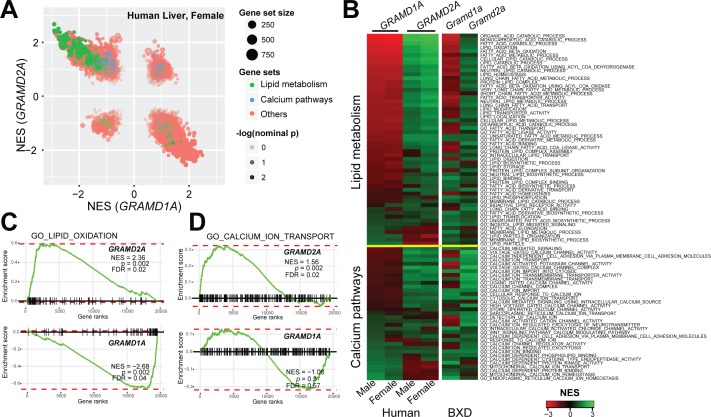
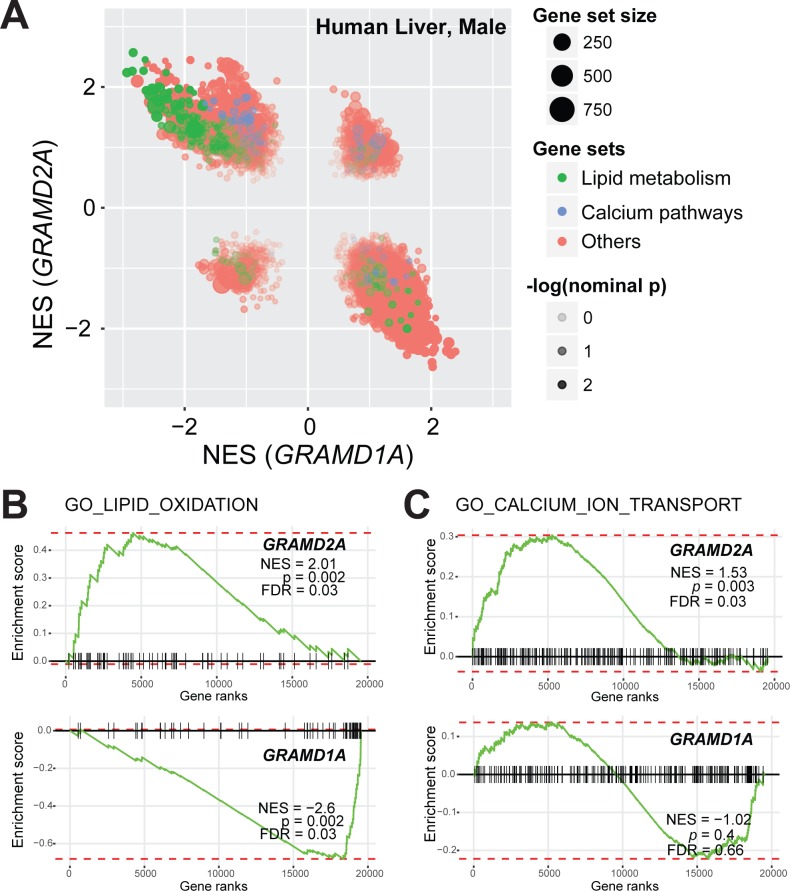
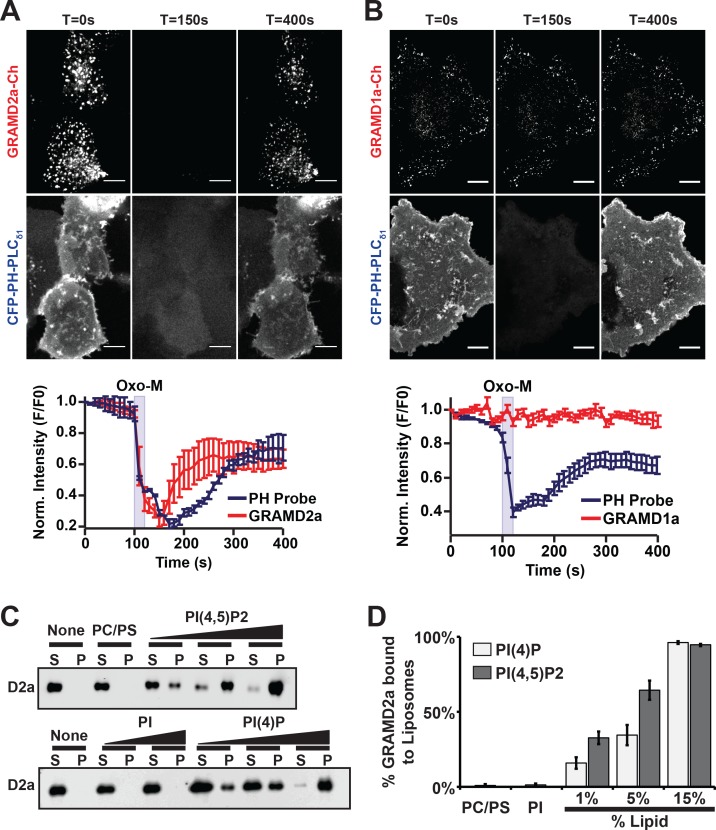
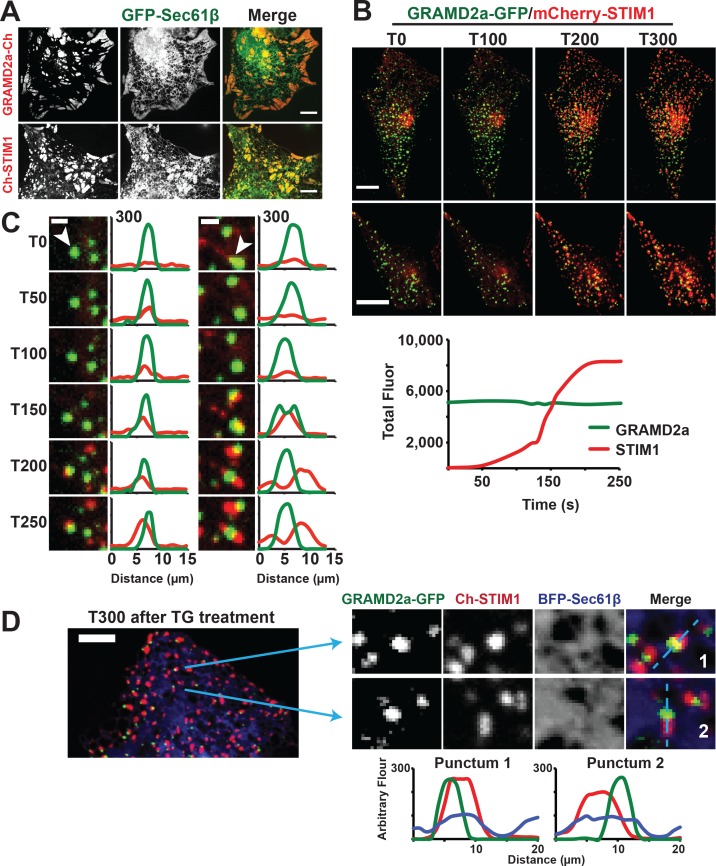
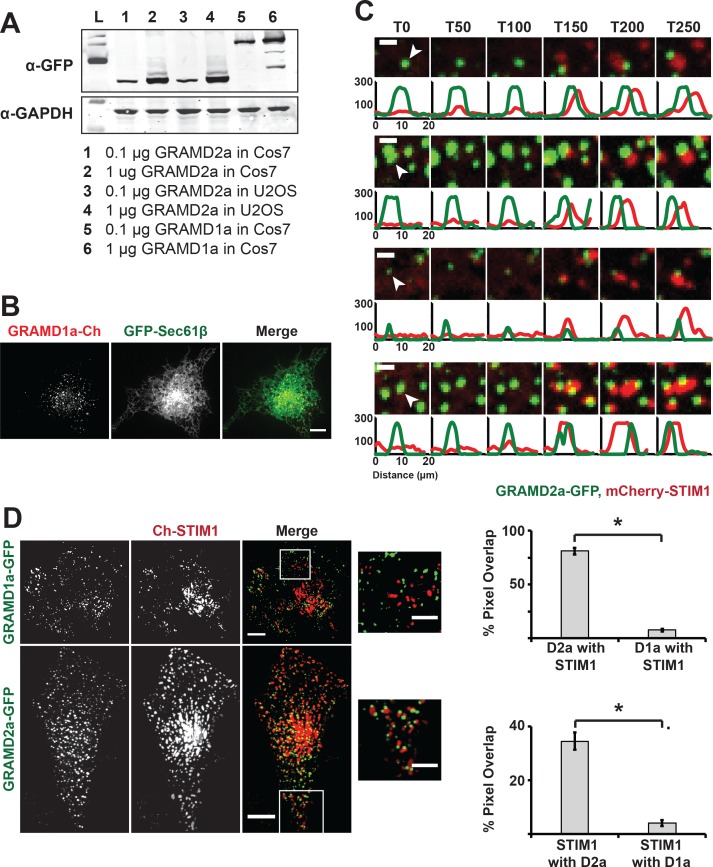
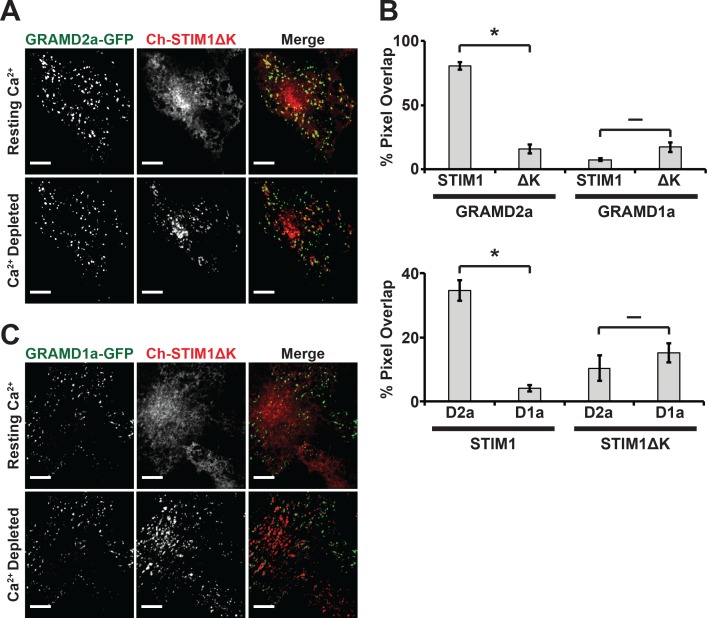
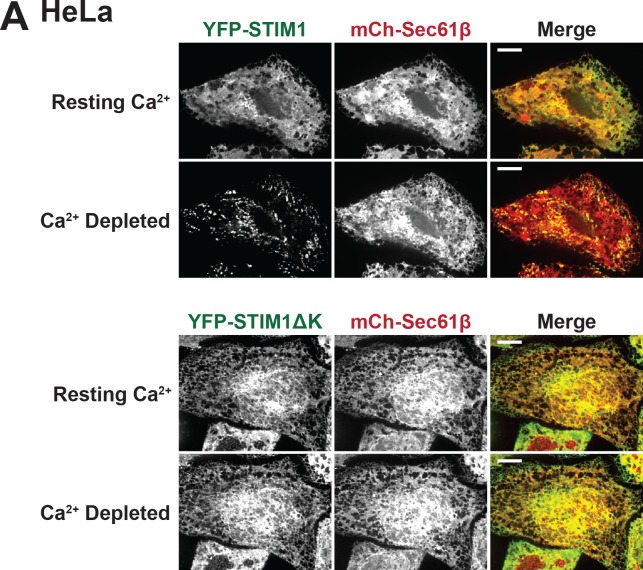
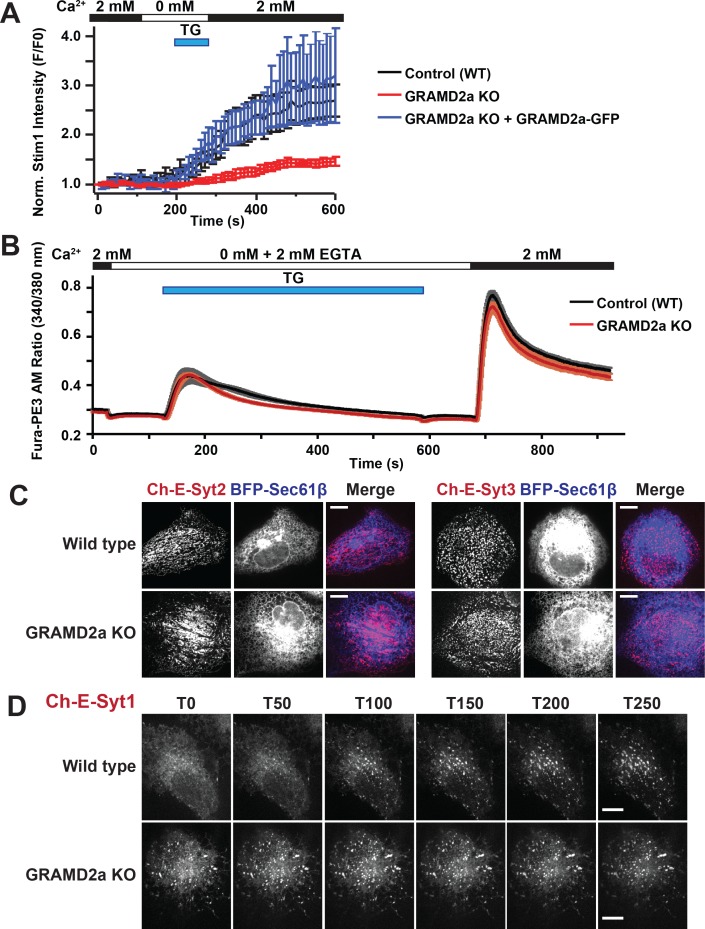
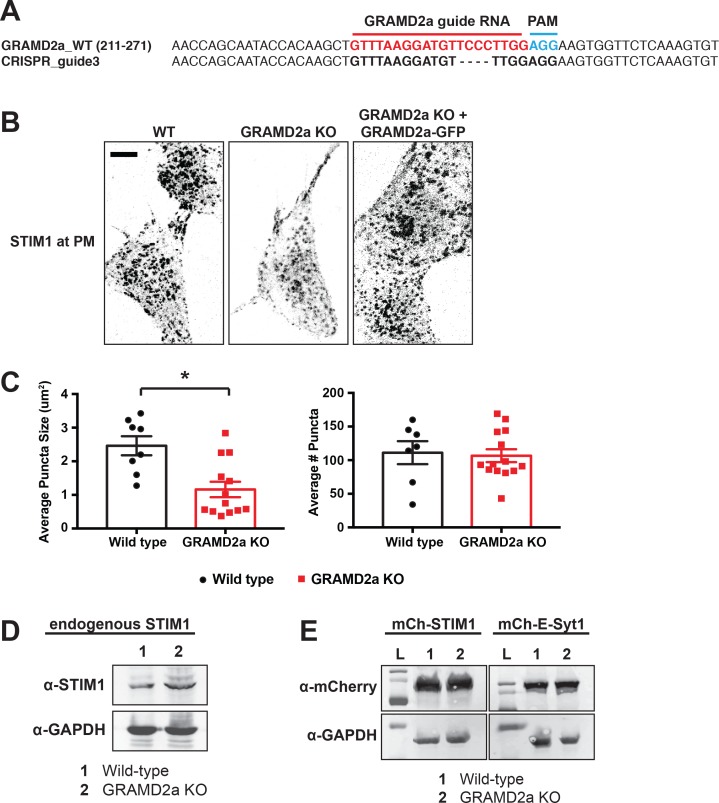
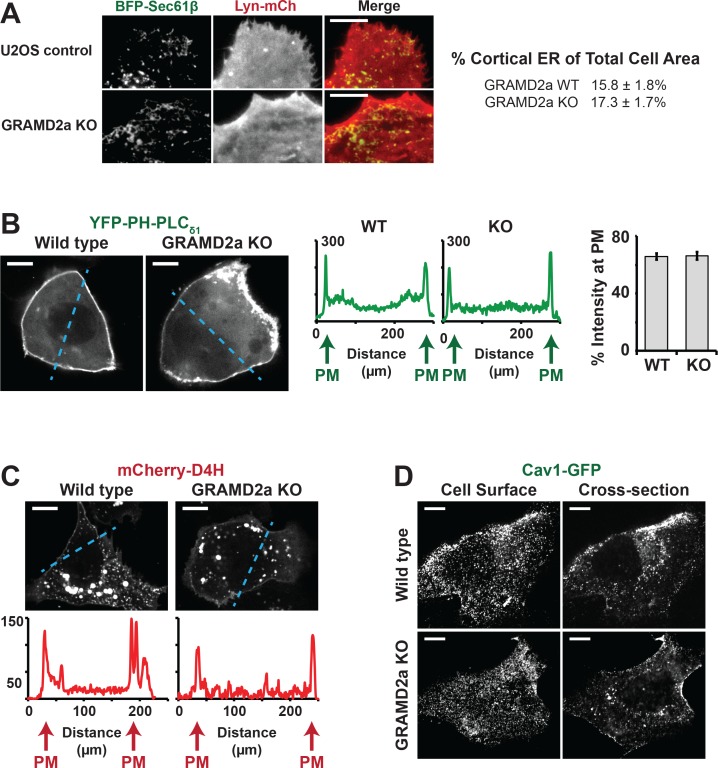
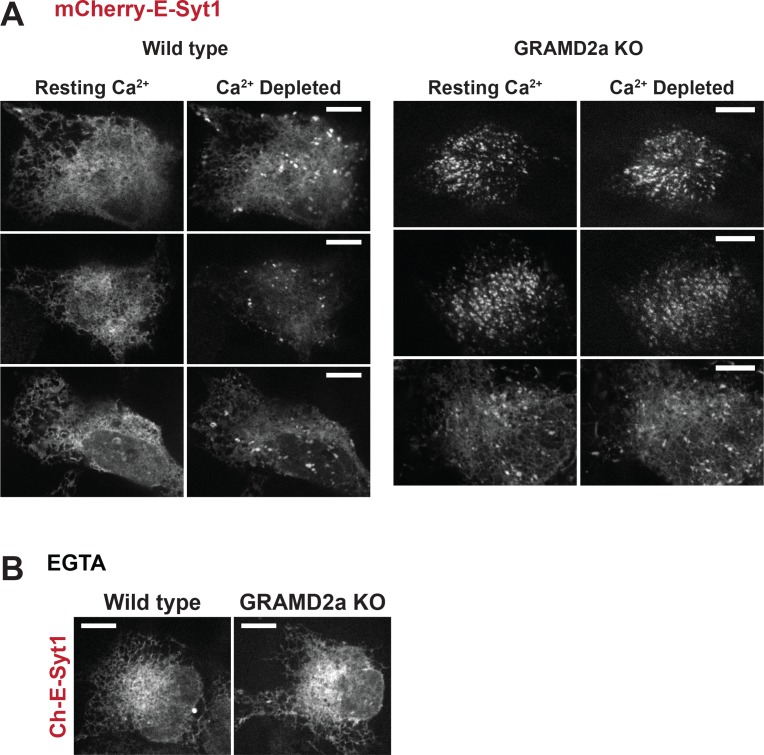
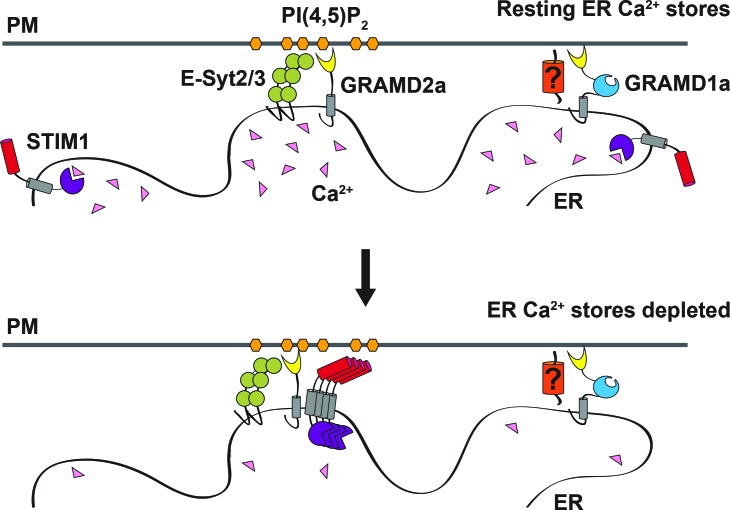
Endoplasmic reticulum (ER) membrane contact sites (MCSs) are crucial regulatory hubs in cells, playing roles in signaling, organelle dynamics, and ion and lipid homeostasis. Previous work demonstrated that the highly conserved yeast Ltc/Lam sterol transporters localize and function at ER MCSs. Our analysis of the human family members, GRAMD1a and GRAMD2a, demonstrates that they are ER-PM MCS proteins, which mark separate regions of the plasma membrane (PM) and perform distinct functions in vivo. GRAMD2a, but not GRAMD1a, co-localizes with the E-Syt2/3 tethers at ER-PM contacts in a PIP lipid-dependent manner and pre-marks the subset of PI(4,5)P2-enriched ER-PM MCSs utilized for STIM1 recruitment. Data from an analysis of cells lacking GRAMD2a suggest that it is an organizer of ER-PM MCSs with pleiotropic functions including calcium homeostasis. Thus, our data demonstrate the existence of multiple ER-PM domains in human cells that are functionally specialized by GRAM-domain containing proteins.
Keywords: ER-PM contact sites; GRAMD proteins; PIP lipids; cell biology; cortical ER; human; membrane contact site; store operated calcium entry.
© 2018, Besprozvannaya et al.
Conflict of interest statement
MB, ED, HL, KG, DB No competing interests declared, JA Scientific advisory board, Mitobridge. JN Jodi Nunnari: Scientific advisory board, Mitobridge.
Figures
Comment in
-
GRAM marks the spot for STIM. Commentary on "GRAM domain proteins specialize functionally distinct ER-PM contact sites in human cells".Cell Calcium. 2018 Jul;73:70-71. doi: 10.1016/j.ceca.2018.04.002. Epub 2018 Apr 18. Cell Calcium. 2018. PMID: 29684786
References
Publication types
MeSH terms
Substances
Grants and funding
- P30 CA093373/CA/NCI NIH HHS/United States
- S10 OD018223/OD/NIH HHS/United States
- R01 AG043930/AG/NIA NIH HHS/United States
- F32GM117689/GM/NIGMS NIH HHS/United States
- R01 HL030077/HL/NHLBI NIH HHS/United States
- R01 AA016957/AA/NIAAA NIH HHS/United States
- R01 GM097432/GM/NIGMS NIH HHS/United States
- R01 GM062942/GM/NIGMS NIH HHS/United States
- R37 GM097432/GM/NIGMS NIH HHS/United States
- R01GM062942/GM/NIGMS NIH HHS/United States
- R01GM097432/GM/NIGMS NIH HHS/United States
- R01 GM106019/GM/NIGMS NIH HHS/United States
- F32 GM117689/GM/NIGMS NIH HHS/United States
- R37 HL030077/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
