

Structural principles that enable oligomeric small heat-shock protein paralogs to evolve distinct functions
- PMID: 29472485
- PMCID: PMC6587588
- DOI: 10.1126/science.aam7229
Structural principles that enable oligomeric small heat-shock protein paralogs to evolve distinct functions
Abstract
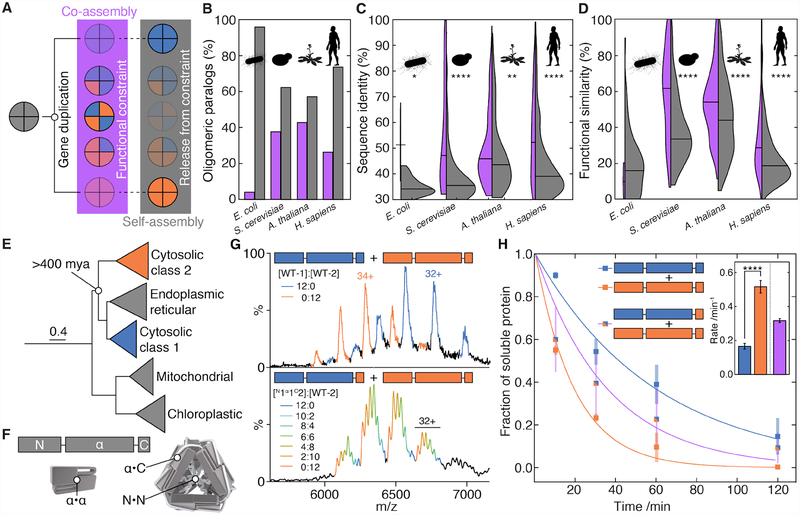
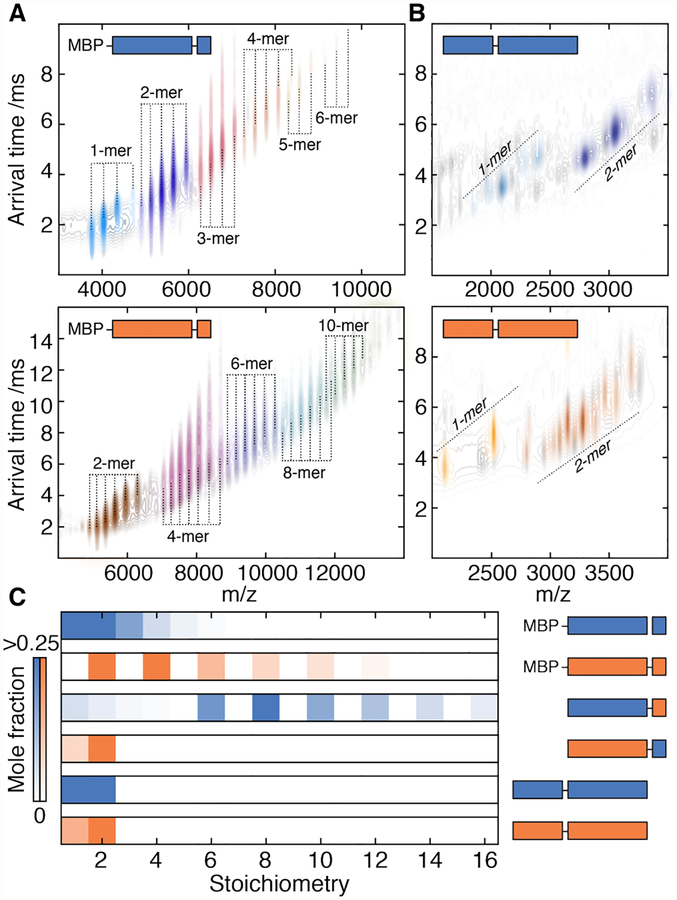
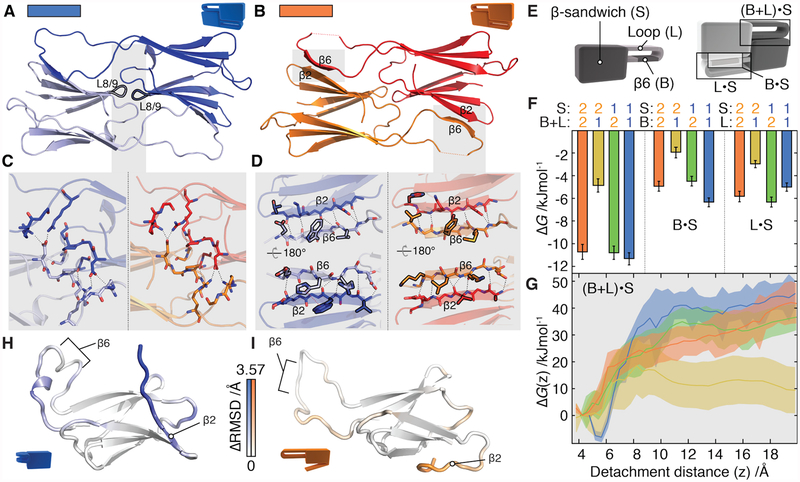
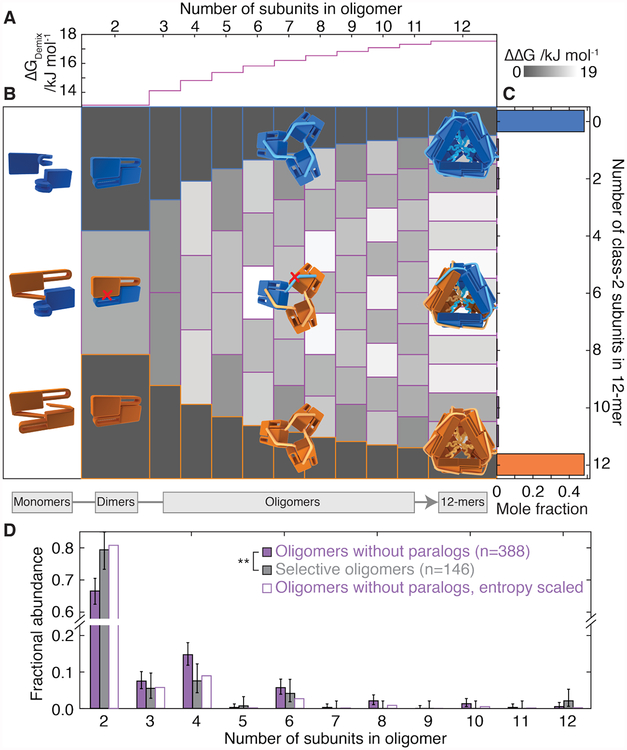
Oligomeric proteins assemble with exceptional selectivity, even in the presence of closely related proteins, to perform their cellular roles. We show that most proteins related by gene duplication of an oligomeric ancestor have evolved to avoid hetero-oligomerization and that this correlates with their acquisition of distinct functions. We report how coassembly is avoided by two oligomeric small heat-shock protein paralogs. A hierarchy of assembly, involving intermediates that are populated only fleetingly at equilibrium, ensures selective oligomerization. Conformational flexibility at noninterfacial regions in the monomers prevents coassembly, allowing interfaces to remain largely conserved. Homomeric oligomers must overcome the entropic benefit of coassembly and, accordingly, homomeric paralogs comprise fewer subunits than homomers that have no paralogs.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Marsh JA, Teichmann SA, Structure, dynamics, assembly, and evolution of protein complexes. Annu Rev Biochem, (2014). - PubMed
-
- Kaltenegger E, Ober D, Paralogue interference affects the dynamics after gene duplication. Trends Plant Sci 20, 814–821 (2015). - PubMed
-
- Diss G et al., Gene duplication can impart fragility, not robustness, in the yeast protein interaction network. Science 355, 630–634 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
