

BigR is a sulfide sensor that regulates a sulfur transferase/dioxygenase required for aerobic respiration of plant bacteria under sulfide stress
- PMID: 29472641
- PMCID: PMC5823870
- DOI: 10.1038/s41598-018-21974-x
BigR is a sulfide sensor that regulates a sulfur transferase/dioxygenase required for aerobic respiration of plant bacteria under sulfide stress
Abstract
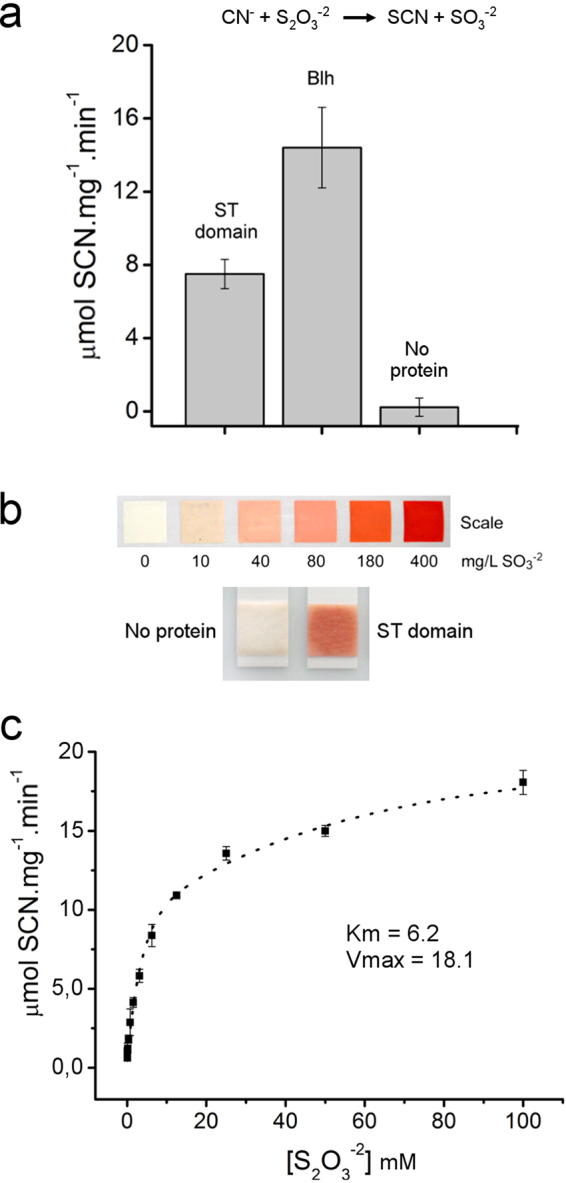
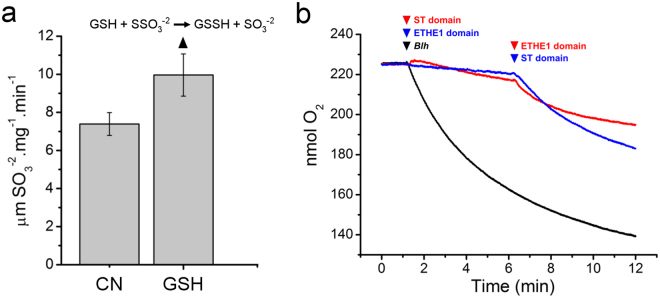
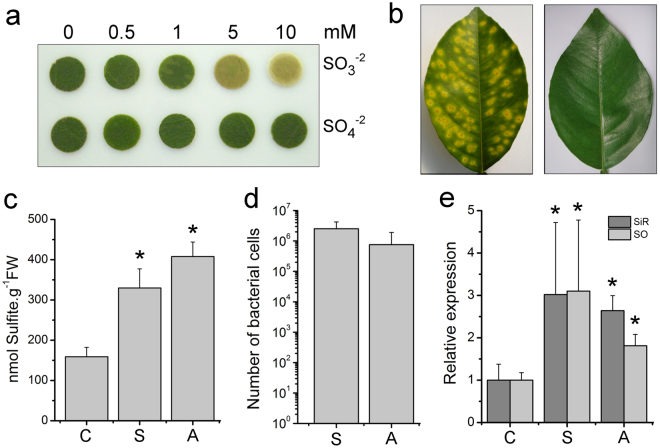
To cope with toxic levels of H2S, the plant pathogens Xylella fastidiosa and Agrobacterium tumefaciens employ the bigR operon to oxidize H2S into sulfite. The bigR operon is regulated by the transcriptional repressor BigR and it encodes a bifunctional sulfur transferase (ST) and sulfur dioxygenase (SDO) enzyme, Blh, required for H2S oxidation and bacterial growth under hypoxia. However, how Blh operates to enhance bacterial survival under hypoxia and how BigR is deactivated to derepress operon transcription is unknown. Here, we show that the ST and SDO activities of Blh are in vitro coupled and necessary to oxidize sulfide into sulfite, and that Blh is critical to maintain the oxygen flux during A. tumefaciens respiration when oxygen becomes limited to cells. We also show that H2S and polysulfides inactivate BigR leading to operon transcription. Moreover, we show that sulfite, which is produced by Blh in the ST and SDO reactions, is toxic to Citrus sinensis and that X. fastidiosa-infected plants accumulate sulfite and higher transcript levels of sulfite detoxification enzymes, suggesting that they are under sulfite stress. These results indicate that BigR acts as a sulfide sensor in the H2S oxidation mechanism that allows pathogens to colonize plant tissues where oxygen is a limiting factor.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
