

Role of Su(Hw) zinc finger 10 and interaction with CP190 and Mod(mdg4) proteins in recruiting the Su(Hw) complex to chromatin sites in Drosophila
- PMID: 29474480
- PMCID: PMC5825117
- DOI: 10.1371/journal.pone.0193497
Role of Su(Hw) zinc finger 10 and interaction with CP190 and Mod(mdg4) proteins in recruiting the Su(Hw) complex to chromatin sites in Drosophila
Abstract
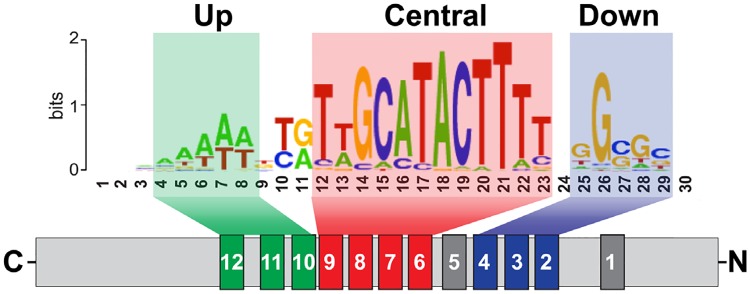
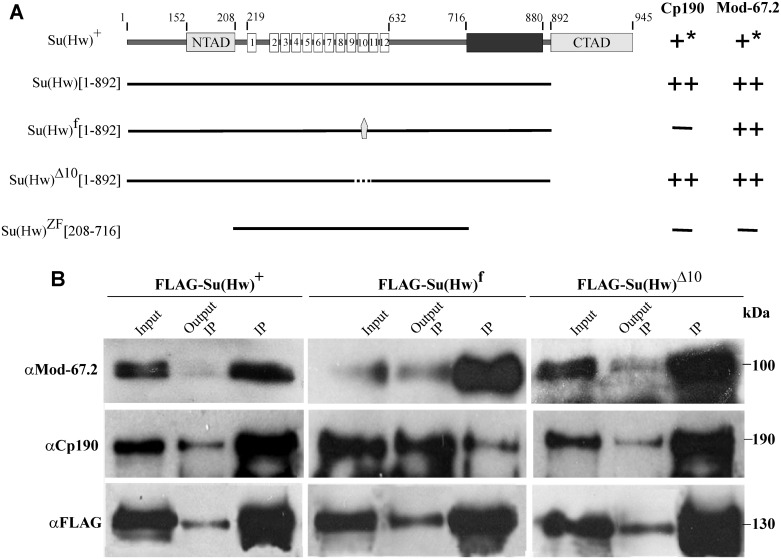
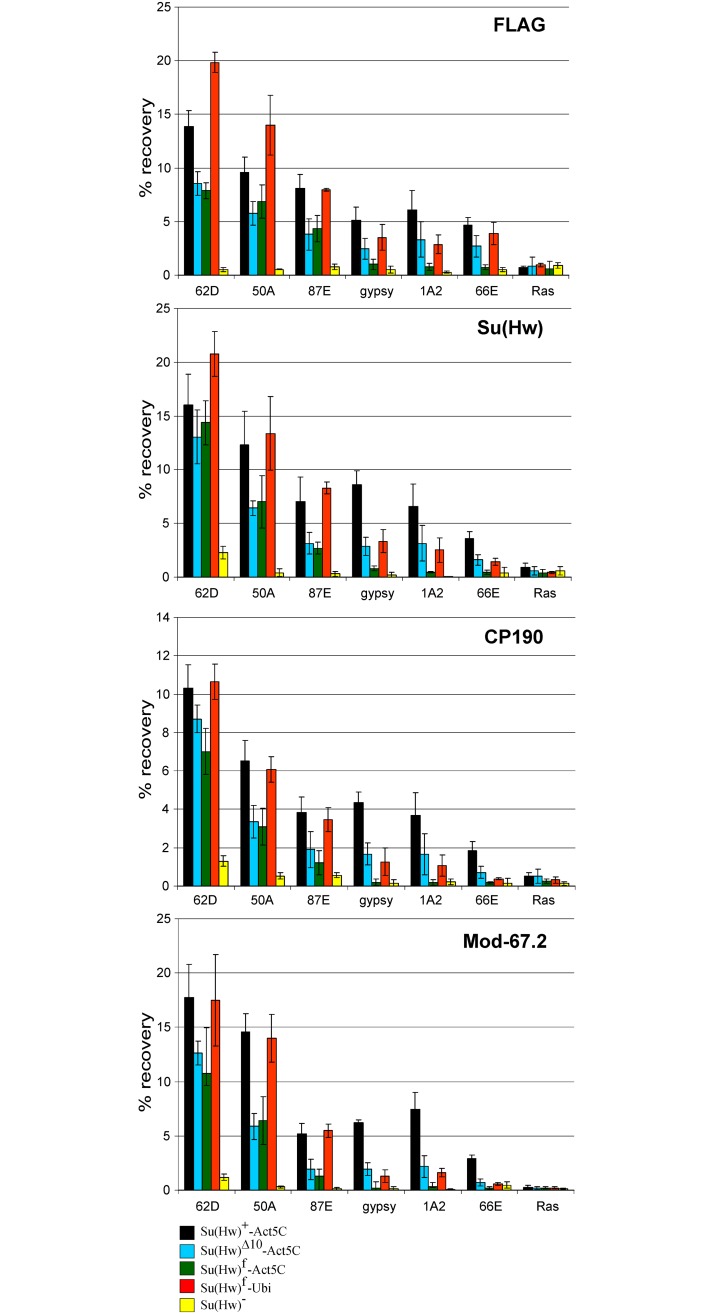
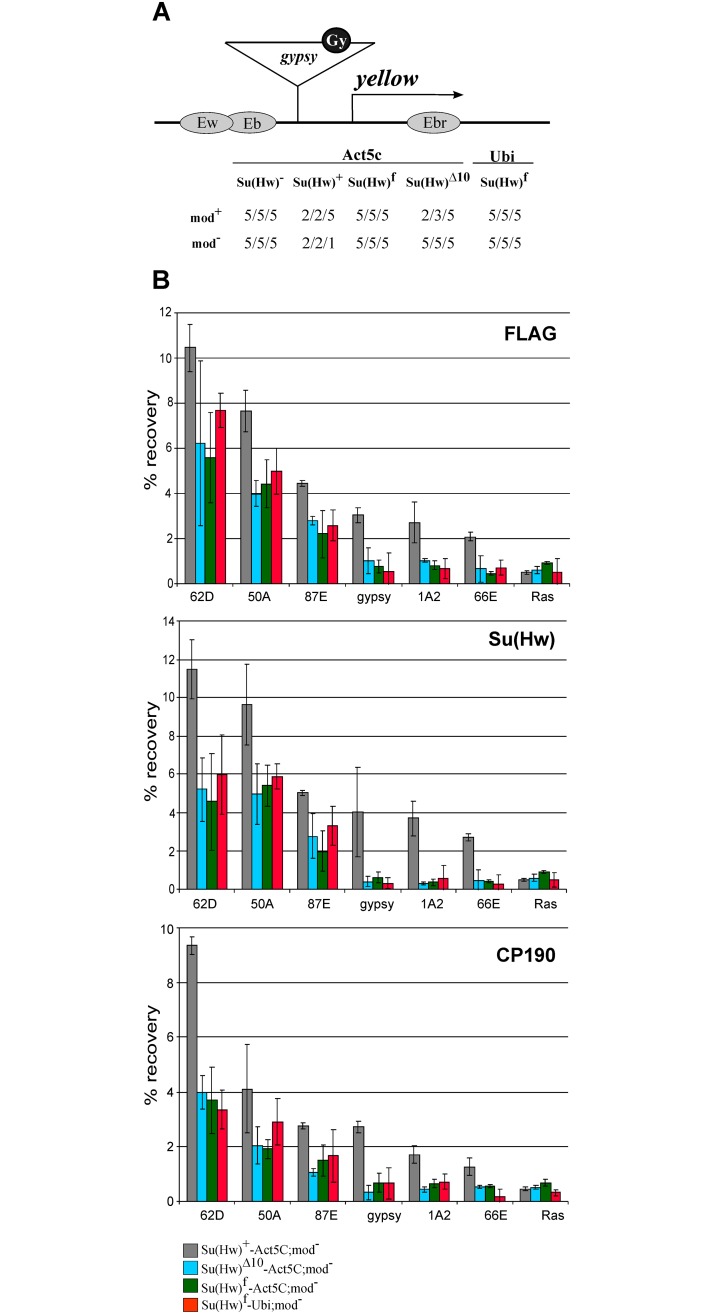
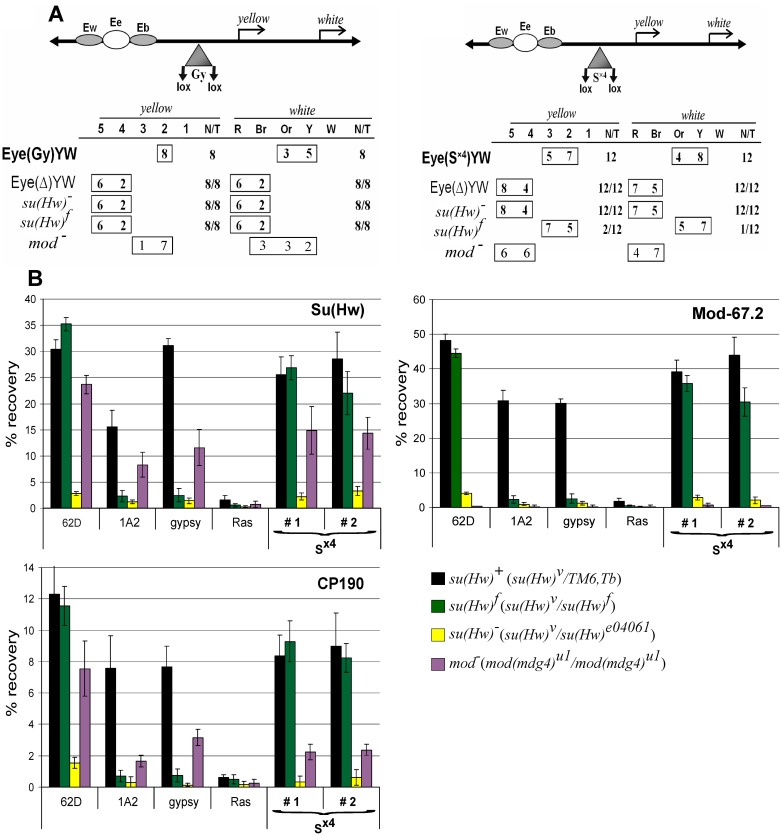
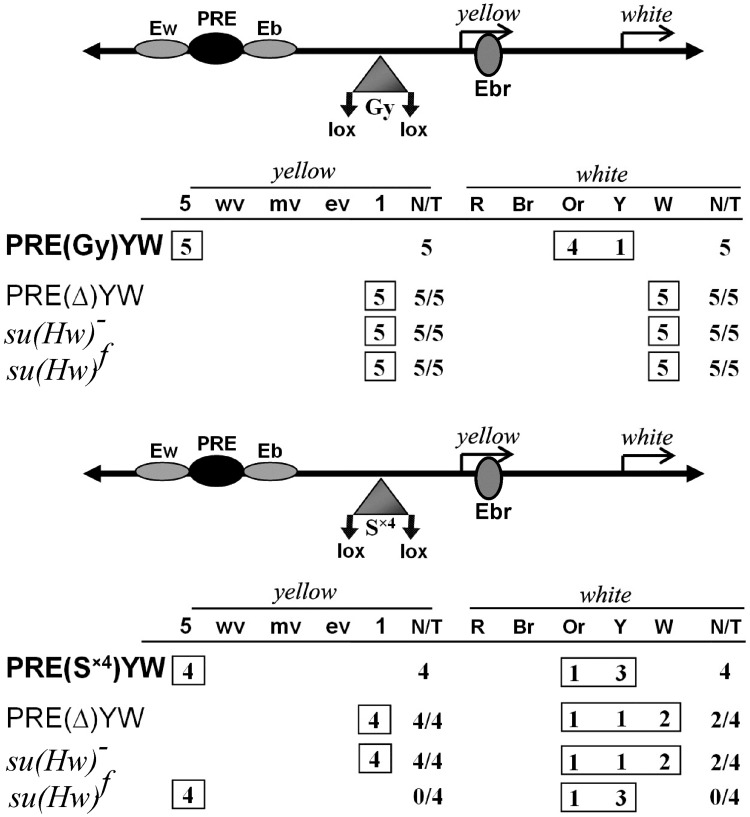
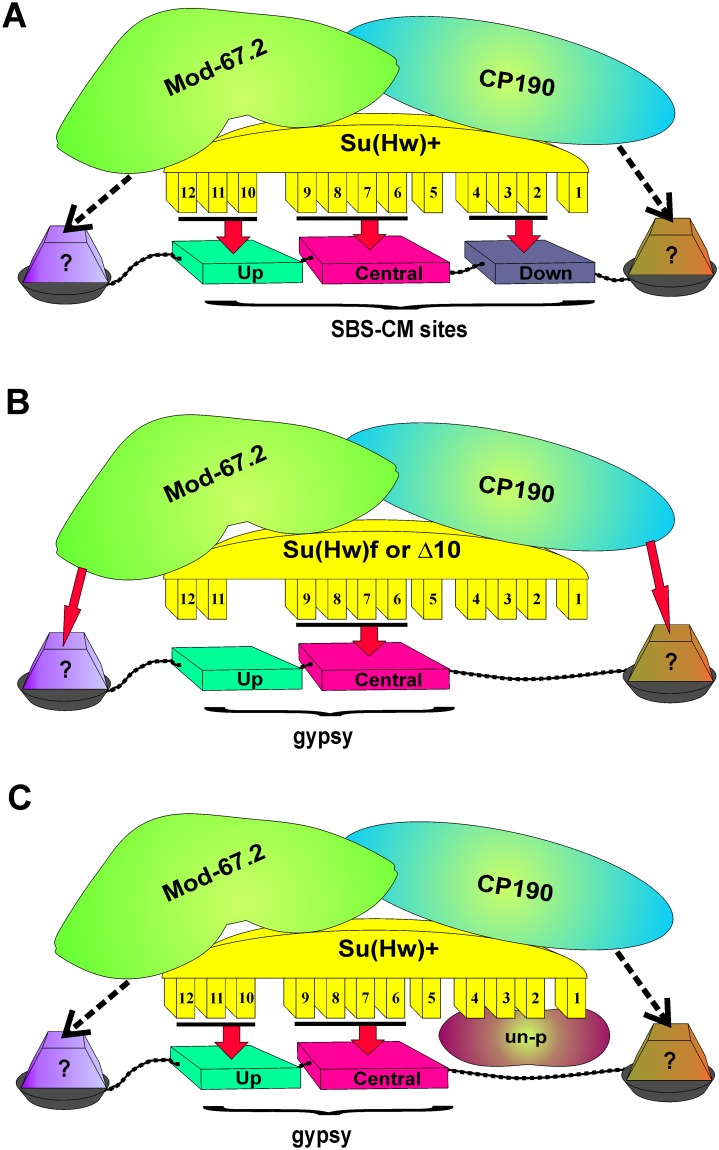
Su(Hw) belongs to the class of proteins that organize chromosome architecture and boundaries/insulators between regulatory domains. This protein contains a cluster of 12 zinc finger domains most of which are responsible for binding to three different modules in the consensus site. Su(Hw) forms a complex with CP190 and Mod(mdg4)-67.2 proteins that binds to well-known Drosophila insulators. To understand how Su(Hw) performs its activities and binds to specific sites in chromatin, we have examined the previously described su(Hw)f mutation that disrupts the 10th zinc finger (ZF10) responsible for Su(Hw) binding to the upstream module. The results have shown that Su(Hw)f loses the ability to interact with CP190 in the absence of DNA. In contrast, complete deletion of ZF10 does not prevent the interaction between Su(Hw)Δ10 and CP190. Having studied insulator complex formation in different mutant backgrounds, we conclude that both association with CP190 and Mod(mdg4)-67.2 partners and proper organization of DNA binding site are essential for the efficient recruitment of the Su(Hw) complex to chromatin insulators.
Conflict of interest statement
Figures
Similar articles
-
Role of Mod(mdg4)-67.2 Protein in Interactions between Su(Hw)-Dependent Complexes and Their Recruitment to Chromatin.Biochemistry (Mosc). 2024 Apr;89(4):626-636. doi: 10.1134/S0006297924040035. Biochemistry (Mosc). 2024. PMID: 38831500
-
Interactions between BTB domain of CP190 and two adjacent regions in Su(Hw) are required for the insulator complex formation.Chromosoma. 2018 Mar;127(1):59-71. doi: 10.1007/s00412-017-0645-6. Epub 2017 Sep 22. Chromosoma. 2018. PMID: 28939920
-
The centrosomal protein CP190 is a component of the gypsy chromatin insulator.Mol Cell. 2004 Dec 3;16(5):737-48. doi: 10.1016/j.molcel.2004.11.004. Mol Cell. 2004. PMID: 15574329
-
Genetic and molecular analysis of the gypsy chromatin insulator of Drosophila.Proc Natl Acad Sci U S A. 1996 Sep 3;93(18):9378-83. doi: 10.1073/pnas.93.18.9378. Proc Natl Acad Sci U S A. 1996. PMID: 8790337 Free PMC article. Review.
-
Functional sub-division of the Drosophila genome via chromatin looping: the emerging importance of CP190.Nucleus. 2013 Mar-Apr;4(2):115-22. doi: 10.4161/nucl.23389. Epub 2013 Jan 18. Nucleus. 2013. PMID: 23333867 Free PMC article. Review.
Cited by
-
Mechanisms of Enhancer-Promoter Interactions in Higher Eukaryotes.Int J Mol Sci. 2021 Jan 12;22(2):671. doi: 10.3390/ijms22020671. Int J Mol Sci. 2021. PMID: 33445415 Free PMC article. Review.
-
The Functions and Mechanisms of Action of Insulators in the Genomes of Higher Eukaryotes.Acta Naturae. 2020 Oct-Dec;12(4):15-33. doi: 10.32607/actanaturae.11144. Acta Naturae. 2020. PMID: 33456975 Free PMC article.
-
Adenosine Receptor and Its Downstream Targets, Mod(mdg4) and Hsp70, Work as a Signaling Pathway Modulating Cytotoxic Damage in Drosophila.Front Cell Dev Biol. 2021 Mar 12;9:651367. doi: 10.3389/fcell.2021.651367. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33777958 Free PMC article.
-
Multi-Scale Organization of the Drosophila melanogaster Genome.Genes (Basel). 2021 May 27;12(6):817. doi: 10.3390/genes12060817. Genes (Basel). 2021. PMID: 34071789 Free PMC article. Review.
-
Investigation of the Developmental Requirements of Drosophila HP1 and Insulator Protein Partner, HIPP1.G3 (Bethesda). 2019 Feb 7;9(2):345-357. doi: 10.1534/g3.118.200705. G3 (Bethesda). 2019. PMID: 30514714 Free PMC article.
References
-
- Lieberman-Aiden E, van Berkum NL, Williams L, Imakaev M, Ragoczy T, Telling A, et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 2009;326: 289–93. doi: 10.1126/science.1181369 - DOI - PMC - PubMed
-
- Dixon JR, Selvaraj S, Yue F, Kim A, Li Y, Shen Y, et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 2012;485: 376–80. doi: 10.1038/nature11082 - DOI - PMC - PubMed
-
- Sexton T, Yaffe E, Kenigsberg E, Bantignies F, Leblanc B, Hoichman M, et al. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 2012;148: 458–72. doi: 10.1016/j.cell.2012.01.010 - DOI - PubMed
-
- Ulianov SV, Khrameeva EE, Gavrilov AA, Flyamer IM, Kos P, Mikhaleva EA, et al. Active chromatin and transcription play a key role in chromosome partitioning into topologically associating domains. Genome Res. 2016;26: 70–84. doi: 10.1101/gr.196006.115 - DOI - PMC - PubMed
-
- Ulianov SV, Tachibana-Konwalski K, Razin SV. Single-cell Hi-C bridges microscopy and genome-wide sequencing approaches to study 3D chromatin organization. Bioessays. 2017. doi: 10.1002/bies.201700104 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
