

Inborn Errors of RNA Lariat Metabolism in Humans with Brainstem Viral Infection
- PMID: 29474921
- PMCID: PMC5886375
- DOI: 10.1016/j.cell.2018.02.019
Inborn Errors of RNA Lariat Metabolism in Humans with Brainstem Viral Infection
Abstract
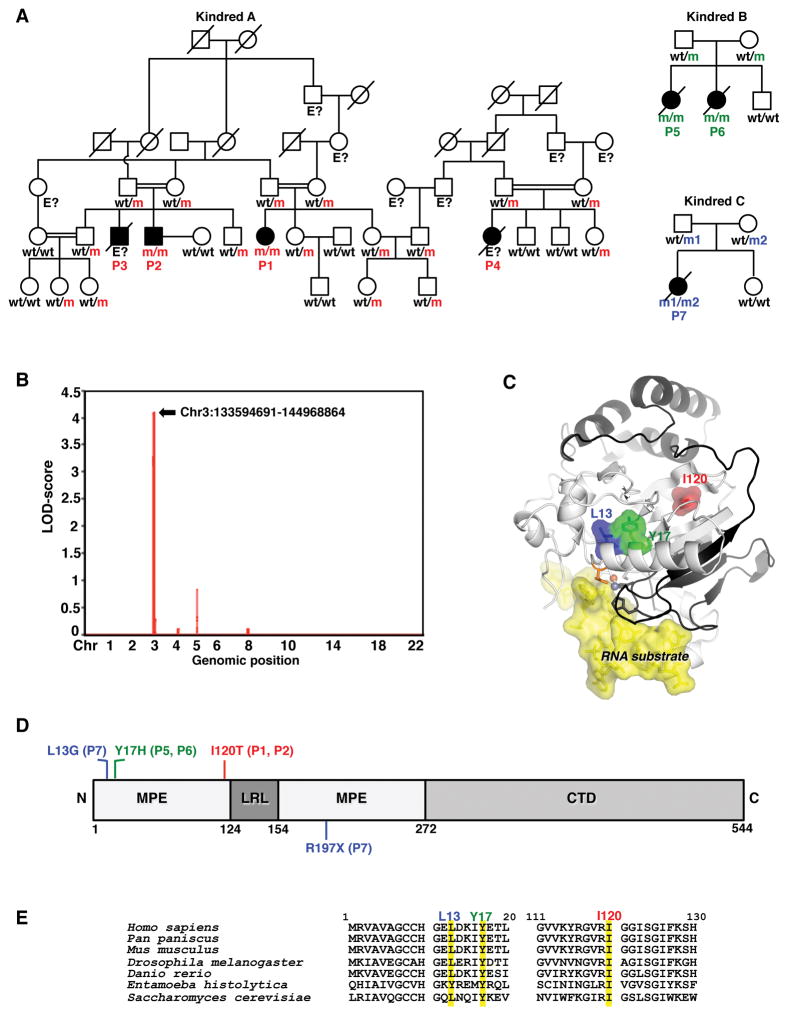
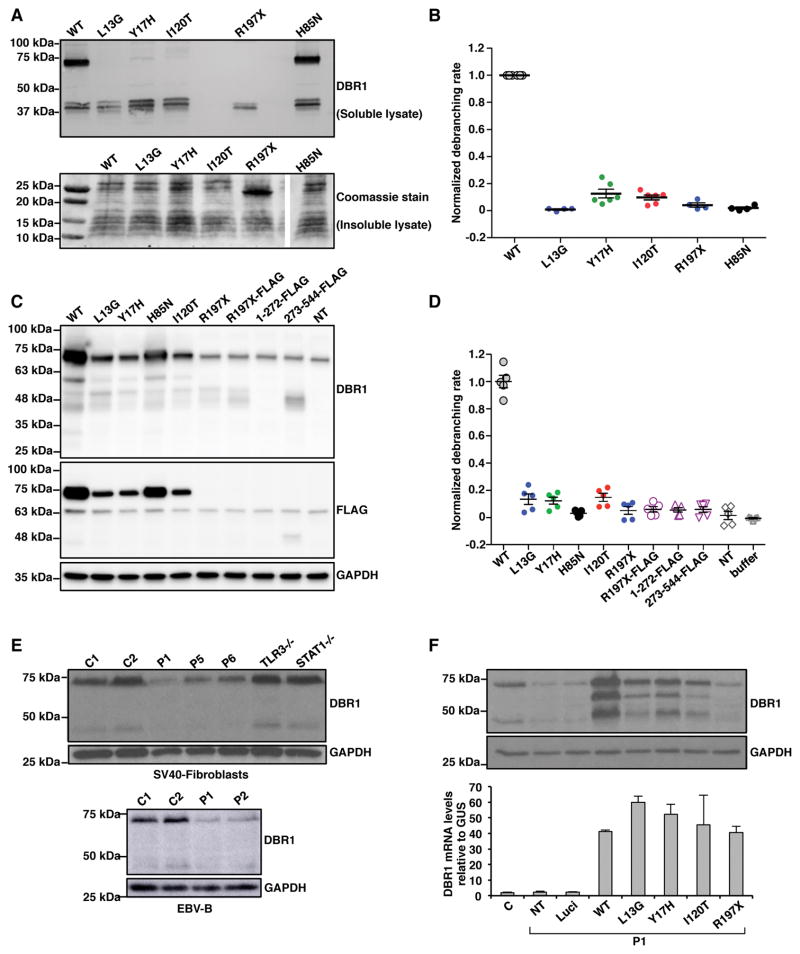
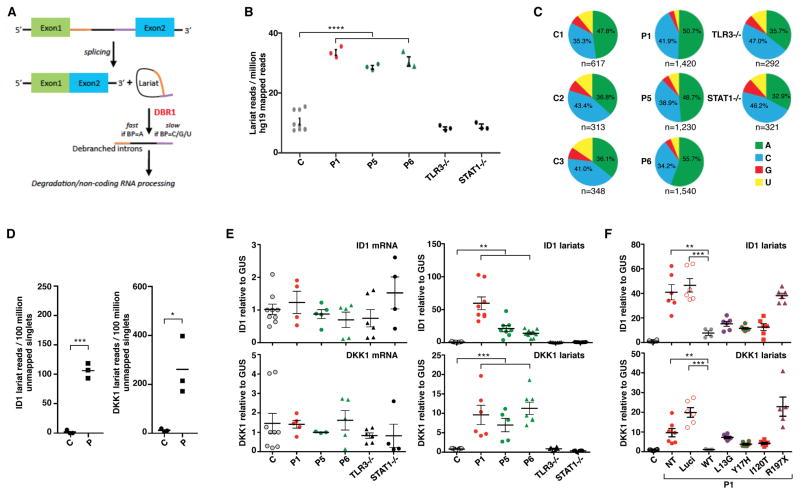
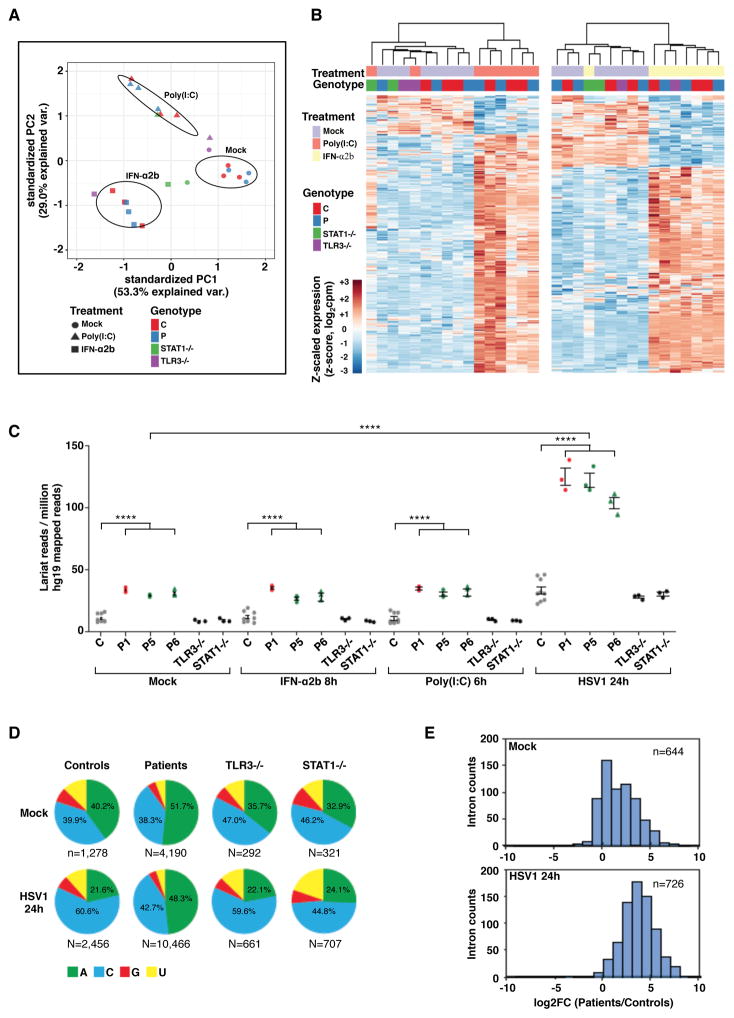
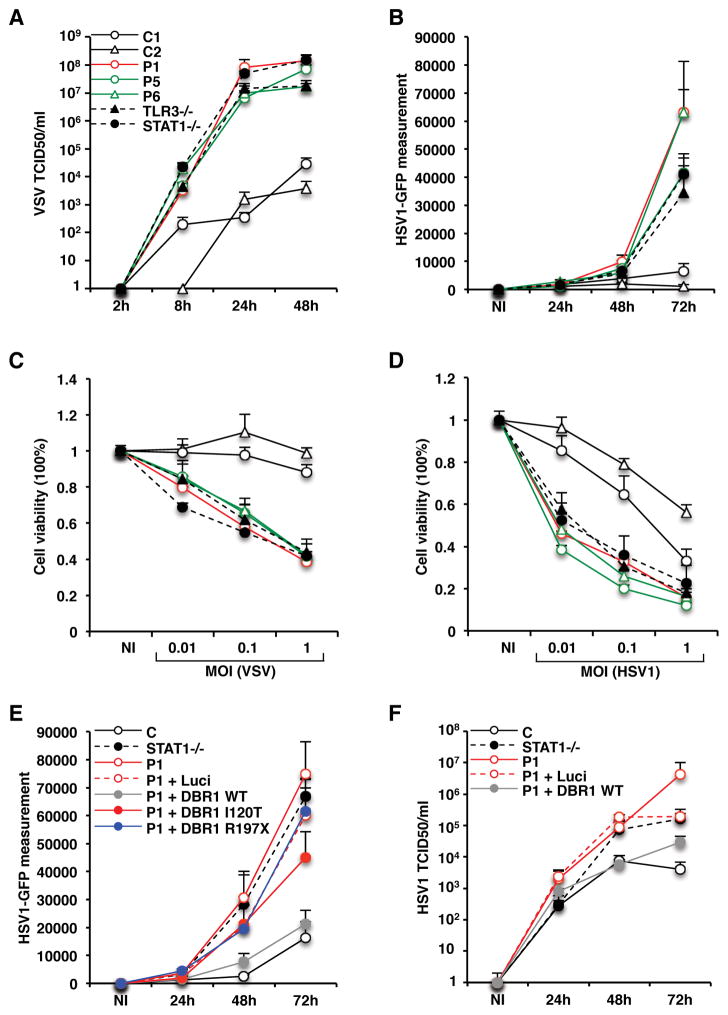
Viruses that are typically benign sometimes invade the brainstem in otherwise healthy children. We report bi-allelic DBR1 mutations in unrelated patients from different ethnicities, each of whom had brainstem infection due to herpes simplex virus 1 (HSV1), influenza virus, or norovirus. DBR1 encodes the only known RNA lariat debranching enzyme. We show that DBR1 expression is ubiquitous, but strongest in the spinal cord and brainstem. We also show that all DBR1 mutant alleles are severely hypomorphic, in terms of expression and function. The fibroblasts of DBR1-mutated patients contain higher RNA lariat levels than control cells, this difference becoming even more marked during HSV1 infection. Finally, we show that the patients' fibroblasts are highly susceptible to HSV1. RNA lariat accumulation and viral susceptibility are rescued by wild-type DBR1. Autosomal recessive, partial DBR1 deficiency underlies viral infection of the brainstem in humans through the disruption of tissue-specific and cell-intrinsic immunity to viruses.
Keywords: DBR1; RNA lariat debranching; brainstem; viral encephalitis.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Abecasis GR, Cherny SS, Cookson WO, Cardon LR. Merlin--rapid analysis of dense genetic maps using sparse gene flow trees. Nat Genet. 2002;30:97–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
