

Single amino acid in V2 encoded by TYLCV is responsible for its self-interaction, aggregates and pathogenicity
- PMID: 29476063
- PMCID: PMC5824789
- DOI: 10.1038/s41598-018-21446-2
Single amino acid in V2 encoded by TYLCV is responsible for its self-interaction, aggregates and pathogenicity
Abstract
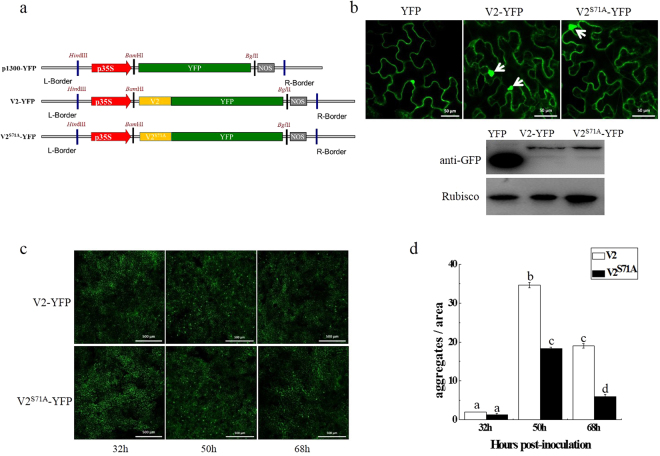
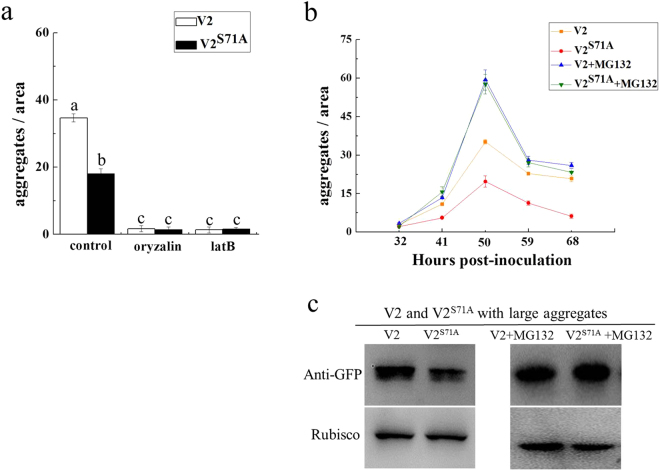
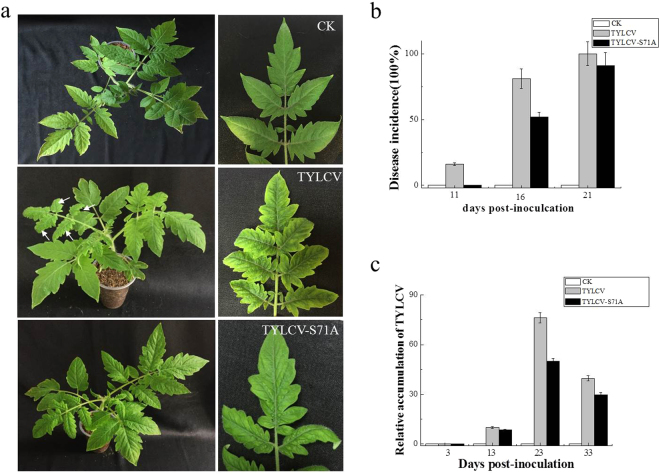
The V2 protein encoded by Begomovirus is essential for virus infection and is involved in multiple functions, such as virus movement and suppression of the host defence response. In this study, we reported that V2 encoded by the Tomato yellow leaf curl virus (TYLCV), which is one of the most devastating tomato-infecting begomoviruses, could interact with itself and a S71A mutation of V2 (V2S71A) abolished its self-interaction. Fluorescence results showed that V2 localized primarily in the cytoplasm and around the nucleus. Site-directed mutagenesis V2S71A had the similar subcellular localization, but V2S71A formed fewer large aggregates in the cytoplasm compared to wild-type V2, whereas the level of aggregates came to a similar after treatment with MG132, which indicates that the S71A mutation might affect 26S proteasome-mediated degradation of V2 aggregates. Meanwhile, heterologous expression of V2S71A from a Potato virus X vector induced mild symptoms compared to wild-type V2, delay of virus infection associated with mild symptoms was observed in plants inoculated with TYLCV-S71A, which indicates that the amino acid on position 71 is also involved in the pathogenicity of V2. To the best of our knowledge, this report is the first to state that the S71A mutation of V2 encoded by TYLCV affects the self-interaction, aggregate formation and pathogenicity of V2.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Nakhla MK, Maxwell DP. Epidemiology and management of tomato yellow leaf curl disease. Plant Virus Dis. Control. 1997;43:565–583.
-
- Glick E, Levy Y, Gafni Y. The viral etiology of Tomato yellow leaf curl disease–a review. Plant Prot. Sci. 2009;45:81–97. doi: 10.17221/26/2009-PPS. - DOI
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
