

Review
doi: 10.1038/s41556-018-0037-z.
Epub 2018 Feb 23.
Cargo recognition and degradation by selective autophagy
Affiliations
- PMID: 29476151
- PMCID: PMC6028034
- DOI: 10.1038/s41556-018-0037-z
Item in Clipboard
Review
Cargo recognition and degradation by selective autophagy
Nat Cell Biol.
2018 Mar.
Erratum in
-
Author Correction: Cargo recognition and degradation by selective autophagy.Nat Cell Biol. 2023 Jul;25(7):1073. doi: 10.1038/s41556-023-01177-x. Nat Cell Biol. 2023. PMID: 37286635 No abstract available.
Abstract
Macroautophagy, initially described as a non-selective nutrient recycling process, is essential for the removal of multiple cellular components. In the past three decades, selective autophagy has been characterized as a highly regulated and specific degradation pathway for removal of unwanted cytosolic components and damaged and/or superfluous organelles. Here, we discuss different types of selective autophagy, emphasizing the role of ligand receptors and scaffold proteins in providing cargo specificity, and highlight unanswered questions in the field.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
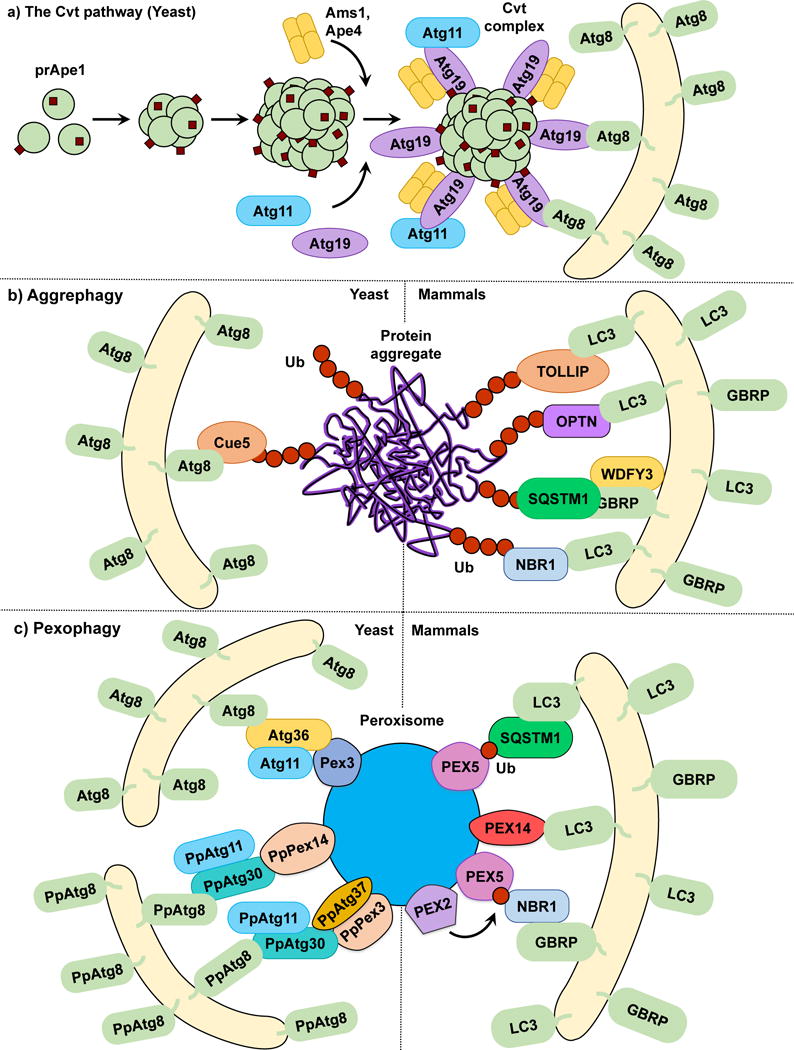
The Cvt pathway, aggrephagy and pexophagy. (a) In the yeast Cvt pathway prApe1, Ape4 and Ams1 are synthesized in the cytoplasm. prApe1 oligomerizes into dodecamers and subsequently higher order structures that are recognized by the receptor Atg19, which in turn binds the scaffold protein Atg11 forming the Cvt Complex. Ams1 and Apr4 also oligomerize and bind Atg19. Atg11 brings the Cvt Complex to the PAS where Atg19 binds Atg8‒PE, tethering the Cvt complex to the phagophore. (b) In both yeast and mammalian aggrephagy, protein aggregates are ubiquitinated and subsequently recognized by cargo receptors. In yeast, Cue5 links the ubiquitinated aggregates to Atg8‒PE. During mammalian aggrephagy, TOLLIP, SQSTM1, NBR1 and OPTN tether the ubiquitinated aggregates to the phagophore by binding LC3/GABARAP family members. WDFY3 has been described as a scaffold for SQSTM1-dependent degradation. (c) In S. cerevisiae pexophagy, Atg36 functions as a receptor linking peroxisomes to the phagophore by binding Pex3 and Atg8‒PE. In P. pastoris pexophagy, PpAtg30 acts as a receptor by linking PpPex3 and PpPex14 to PpAtg8‒PE. Atg11 functions as a scaffold for both S. cerevisiae and P. pastoris. The current model of mammalian pexophagy involves the E3-ubiquitin ligase PEX2-mediated mono-ubiquitination of PEX5, which in turn is recognized by receptors SQSTM1 and NBR1, tethering peroxisomes to the phagophore. PEX14 has also been reported to link peroxisomes to the phagophore by directly binding LC3 family members.
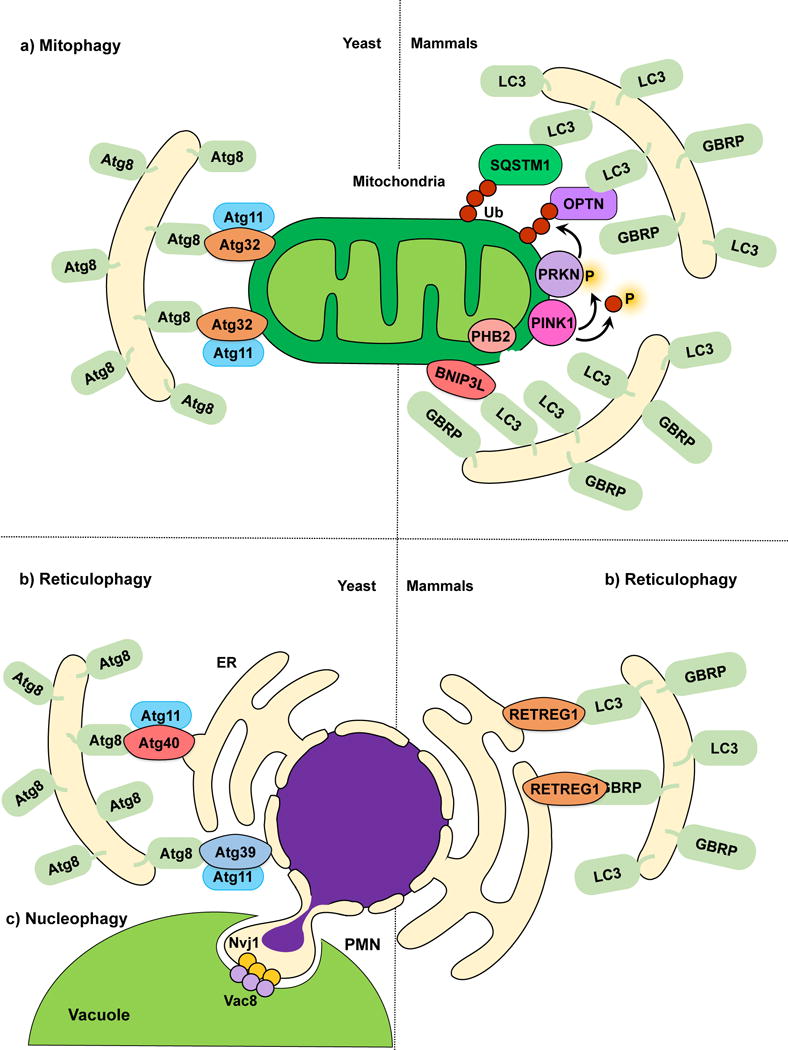
Mitophagy, reticulophagy and nucleophagy. (a) The yeast mitophagy receptor Atg32 links mitochondria to the phagophore by directly binding Atg8‒PE; Atg11 functions as a scaffold. Several cargo receptors (not all shown) have been described for mammalian mitophagy. Mitochondria depolarization leads to PINK1 activation and phosphorylation of ubiquitin and PRKN, and OMM disruption exposes PHB2. Receptors link mitochondria targeted for degradation to the phagophore. (b) In yeast reticulophagy, Atg39 and Atg40 have been proposed as receptor proteins. Atg39 mediates the degradation of the perinuclear ER, and Atg40 mediates cytoplasmic ER degradation. Both Atg39 and Atg40 link their respective ER sites to Atg8‒PE-conjugated membranes for sequestration. Atg11 has been proposed as a scaffold protein for both Atg39 and Atg40-mediated reticulophagy. During mammalian reticulophagy, RETREG1/FAM134B tethers the cytoplasmic ER to LC3/GABARAP family members for membrane sequestration and degradation. (c) Because Atg39 specifically localizes to the perinuclear ER, Atg39-mediated degradation is also considered nucleophagy. During piecemeal microautophagy of the nucleus (PMN), the nuclear envelope protein Nvj1 and vacuolar membrane protein Vac8 form nuclear-vacuolar junctions, which pinch off and engulf part of the nucleus inside the vacuole.
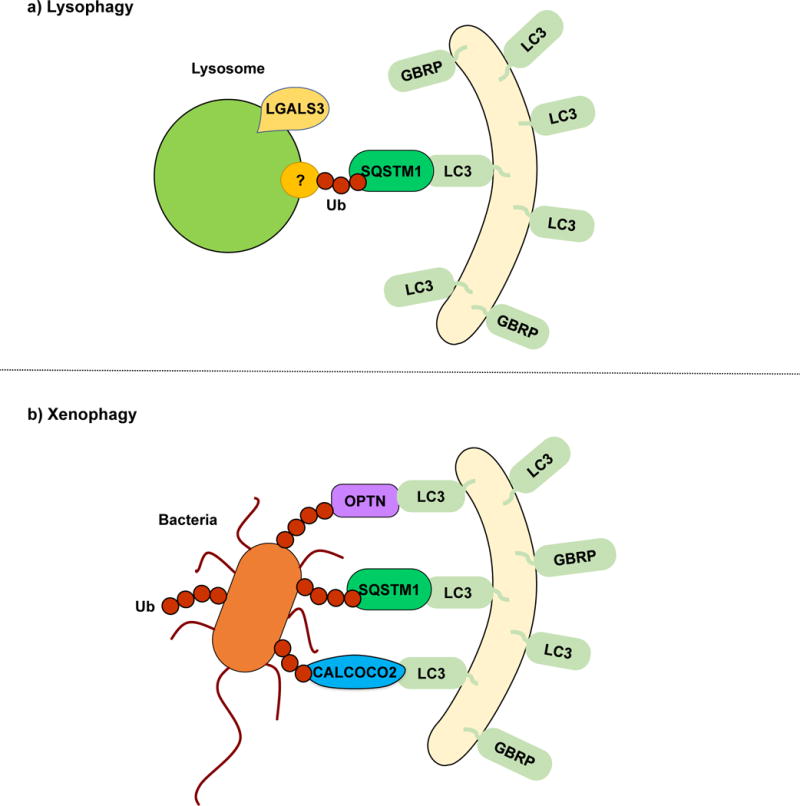
Lysophagy and xenophagy. (a) During lysophagy, unknown lysosomal proteins are ubiquitinated and recognized by SQSTM1, which functions as a receptor, linking the damaged lysosomes with the LC3/GABARAP-conjugated sequestering membranes. LGALS3 binds to exposed lysosomal glycoproteins upon membrane rupture. A specific lysophagy mechanism remains to be elucidated. (b) In xenophagy, intracellular pathogens such as viruses and bacteria are recognized and ubiquitinated. SQSTM1, OPTN, CALCOCO2 and NBR1 have been described as receptor proteins.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
