

Foxm1 controls a pro-stemness microRNA network in neural stem cells
- PMID: 29476172
- PMCID: PMC5824884
- DOI: 10.1038/s41598-018-21876-y
Foxm1 controls a pro-stemness microRNA network in neural stem cells
Abstract
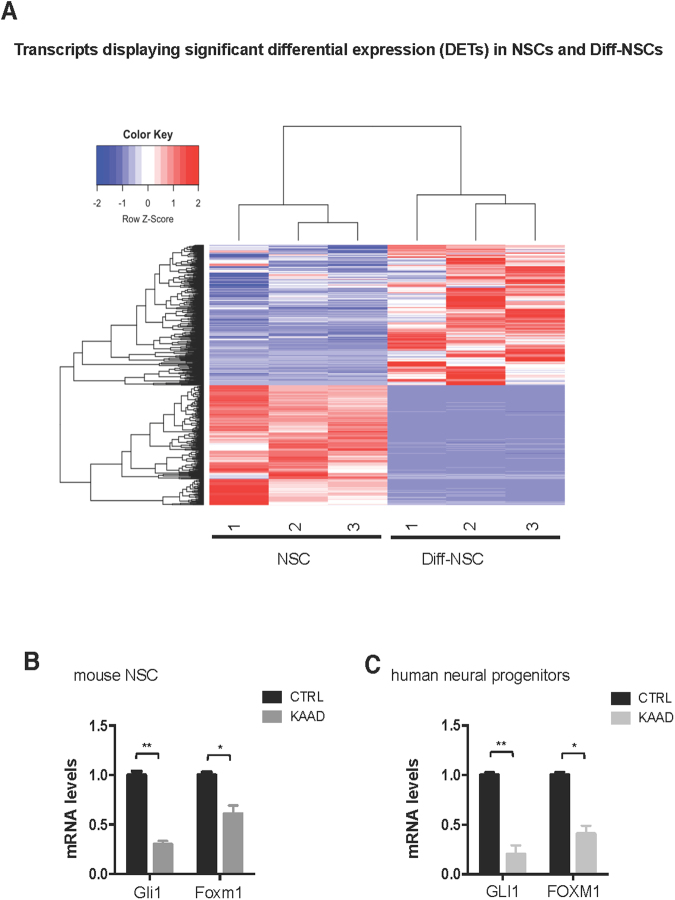
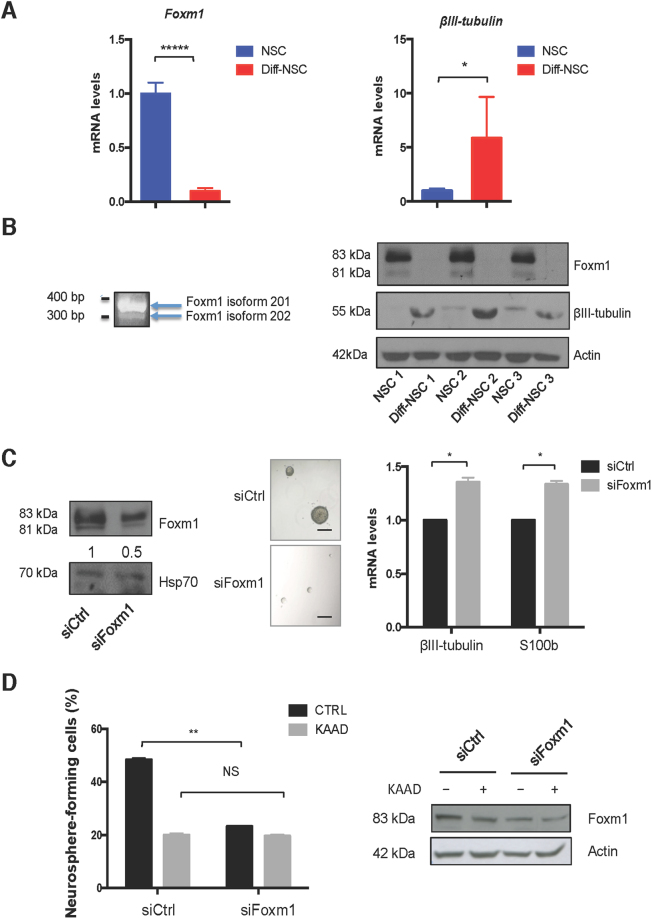
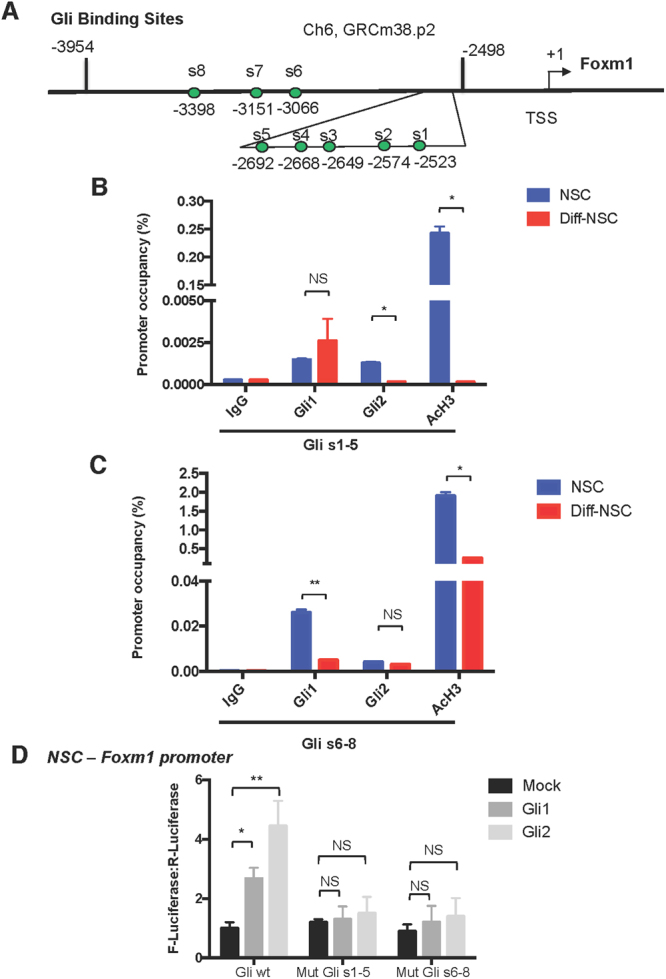
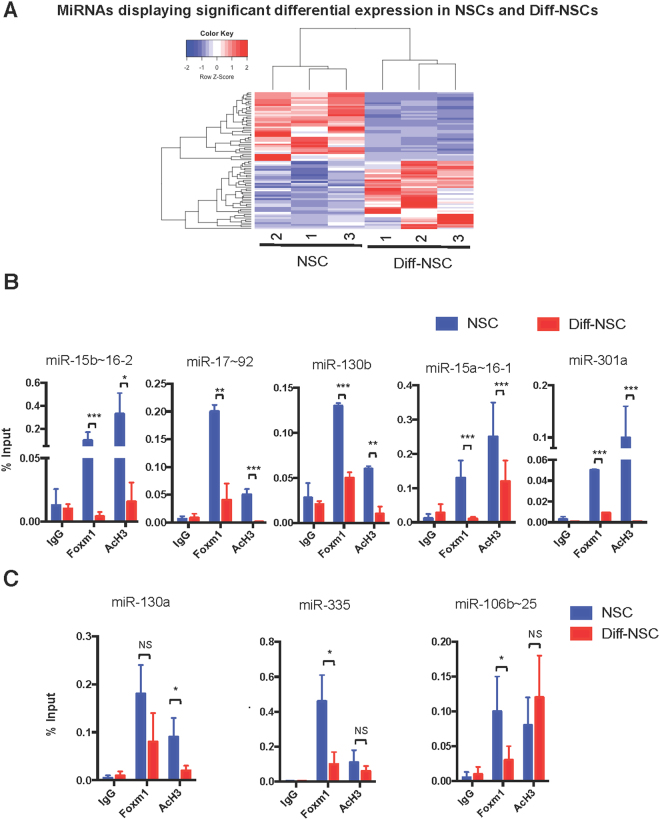
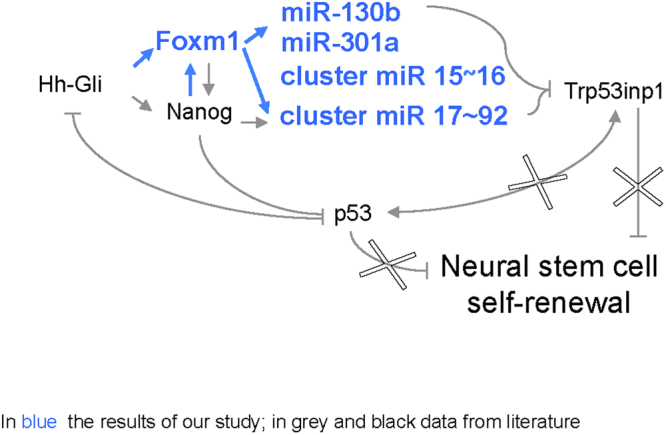
Cerebellar neural stem cells (NSCs) require Hedgehog-Gli (Hh-Gli) signalling for their maintenance and Nanog expression for their self-renewal. To identify novel molecular features of this regulatory pathway, we used next-generation sequencing technology to profile mRNA and microRNA expression in cerebellar NSCs, before and after induced differentiation (Diff-NSCs). Genes with higher transcript levels in NSCs (vs. Diff-NSCs) included Foxm1, which proved to be directly regulated by Gli and Nanog. Foxm1 in turn regulated several microRNAs that were overexpressed in NSCs: miR-130b, miR-301a, and members of the miR-15~16 and miR-17~92 clusters and whose knockdown significantly impaired the neurosphere formation ability. Our results reveal a novel Hh-Gli-Nanog-driven Foxm1-microRNA network that controls the self-renewal capacity of NSCs.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
