

Toll-like receptor 9 antagonizes antibody affinity maturation
- PMID: 29476183
- PMCID: PMC5839995
- DOI: 10.1038/s41590-018-0052-z
Toll-like receptor 9 antagonizes antibody affinity maturation
Abstract
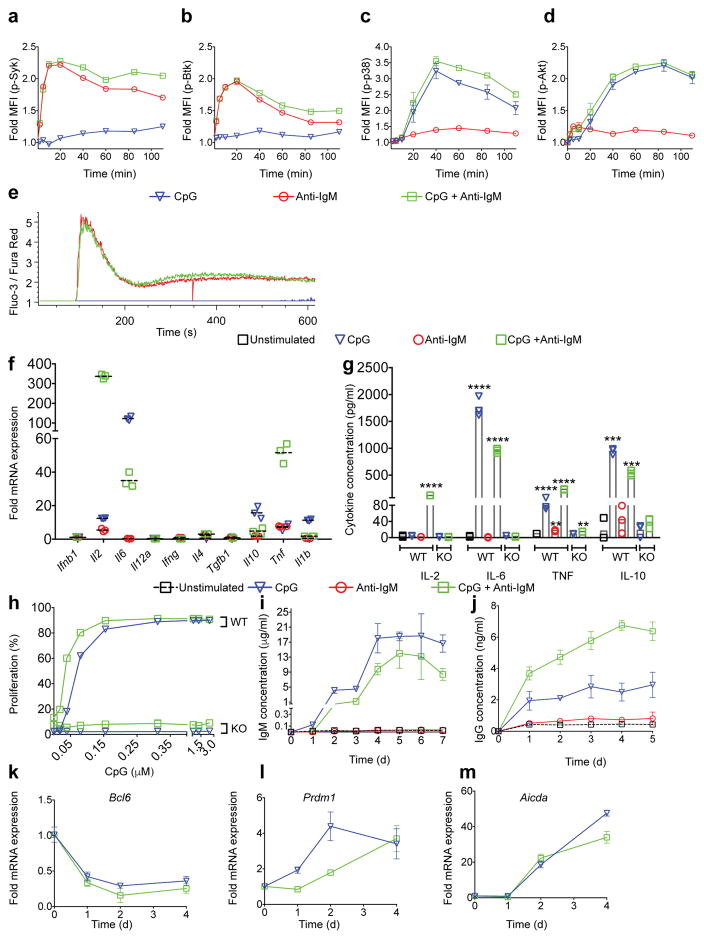
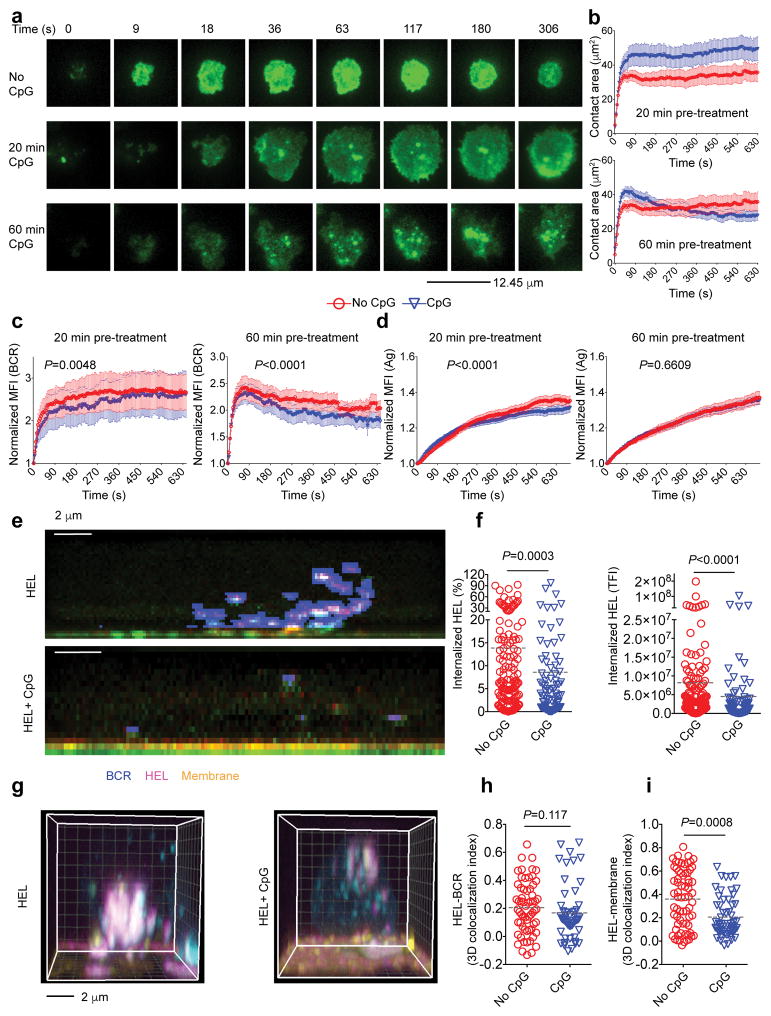
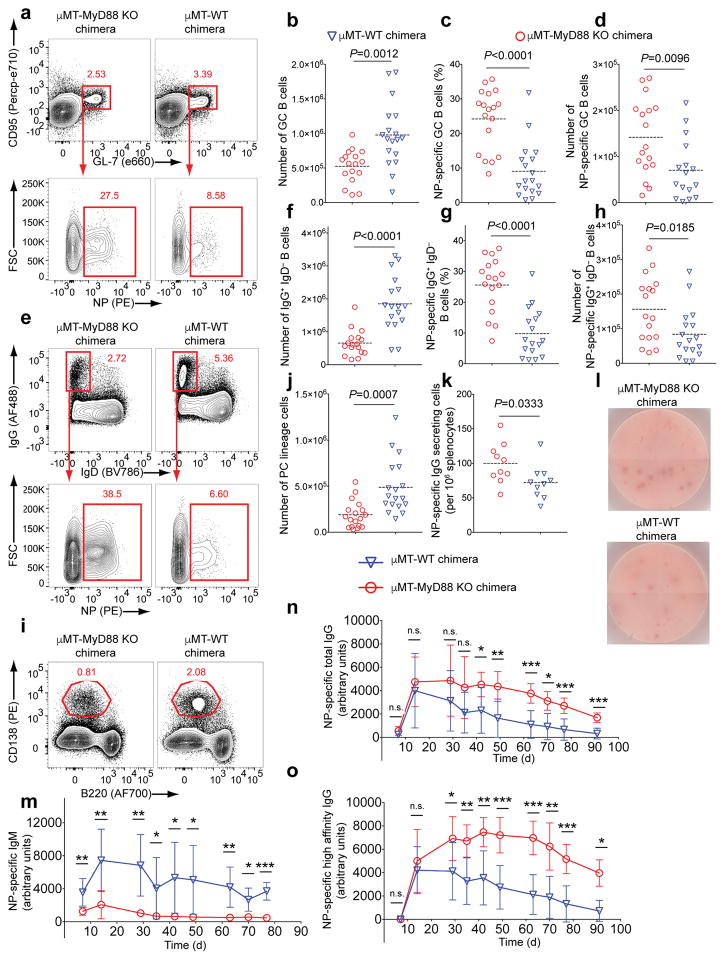
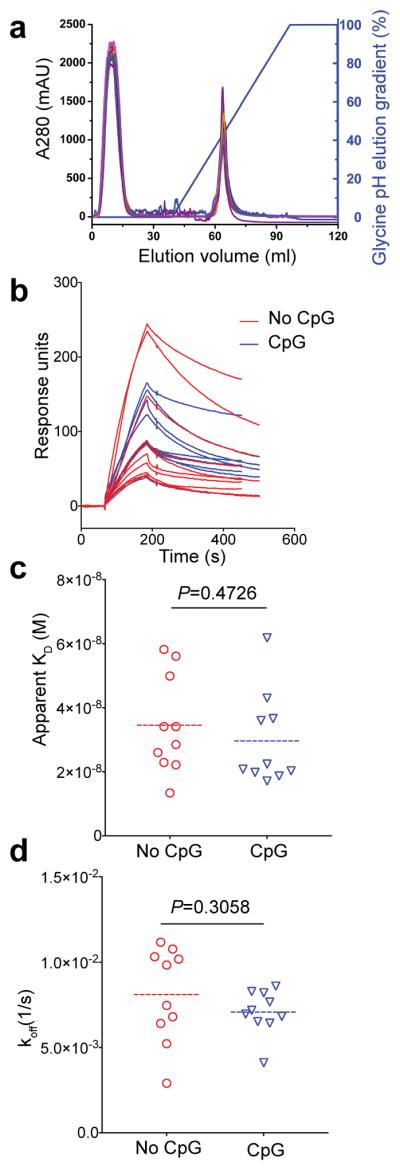
Key events in T cell-dependent antibody responses, including affinity maturation, are dependent on the B cell's presentation of antigen to helper T cells at critical checkpoints in germinal-center formation in secondary lymphoid organs. Here we found that signaling via Toll-like receptor 9 (TLR9) blocked the ability of antigen-specific B cells to capture, process and present antigen and to activate antigen-specific helper T cells in vitro. In a mouse model in vivo and in a human clinical trial, the TLR9 agonist CpG enhanced the magnitude of the antibody response to a protein vaccine but failed to promote affinity maturation. Thus, TLR9 signaling might enhance antibody titers at the expense of the ability of B cells to engage in germinal-center events that are highly dependent on B cells' capture and presentation of antigen.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
