

Exploring the Viral Channel KcvPBCV-1 Function via Computation
- PMID: 29476260
- PMCID: PMC6028866
- DOI: 10.1007/s00232-018-0022-2
Exploring the Viral Channel KcvPBCV-1 Function via Computation
Abstract
Viral potassium channels (Kcv) are homologous to the pore module of complex [Formula: see text]-selective ion channels of cellular organisms. Due to their relative simplicity, they have attracted interest towards understanding the principles of [Formula: see text] conduction and channel gating. In this work, we construct a homology model of the [Formula: see text] open state, which we validate by studying the binding of known blockers and by monitoring ion conduction through the channel. Molecular dynamics simulations of this model reveal that the re-orientation of selectivity filter carbonyl groups coincides with the transport of potassium ions, suggesting a possible mechanism for fast gating. In addition, we show that the voltage sensitivity of this mechanism can originate from the relocation of potassium ions inside the selectivity filter. We also explore the interaction of [Formula: see text] with the surrounding bilayer and observe the binding of lipids in the area between two adjacent subunits. The model is available to the scientific community to further explore the structure/function relationship of Kcv channels.
Keywords: Conduction; Gating; Homology modeling; Molecular dynamics simulations; Protein–lipid interaction; Viral ion channel.
Conflict of interest statement
A. E. V. Andersson, M. A. Kasimova, and L. Delemotte declare that they have no conflict of interest.
Figures
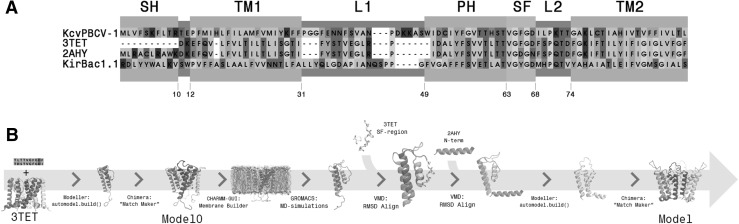
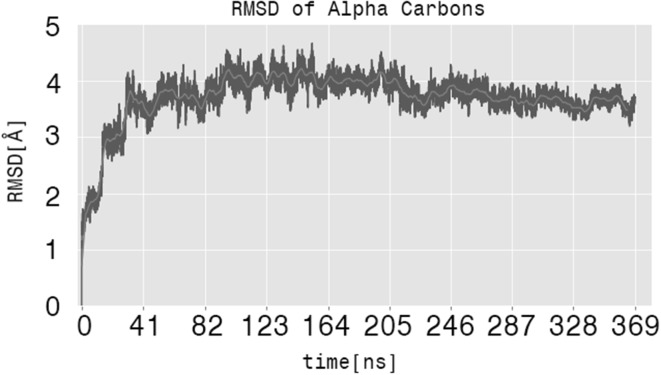
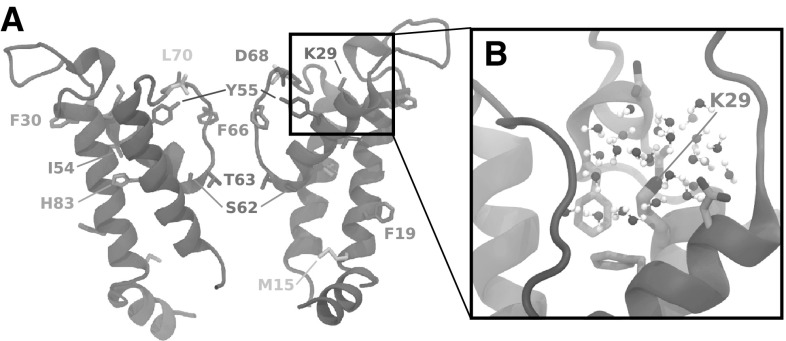
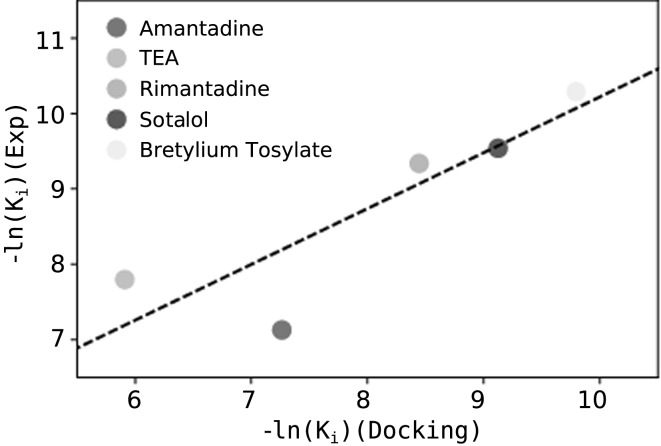
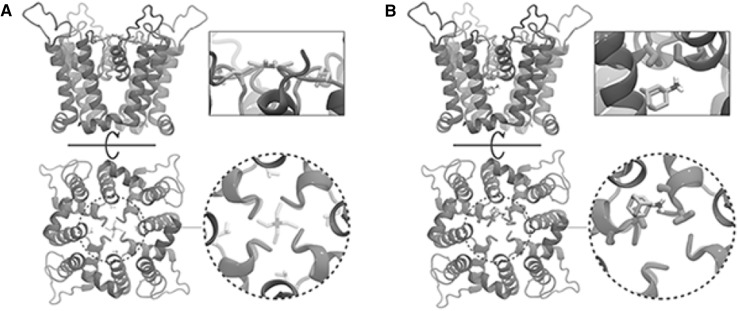
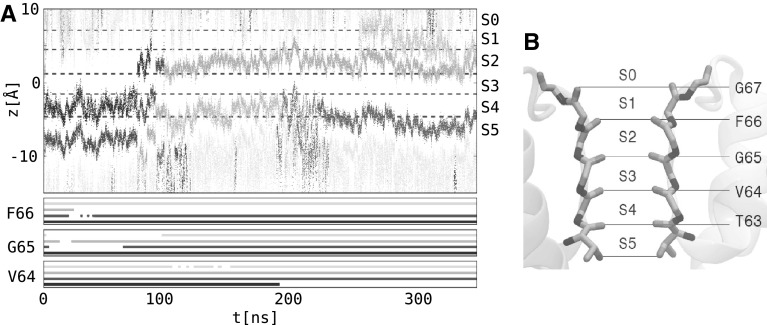


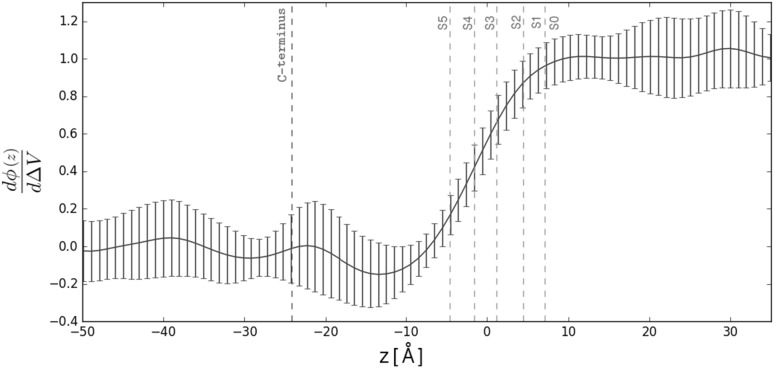
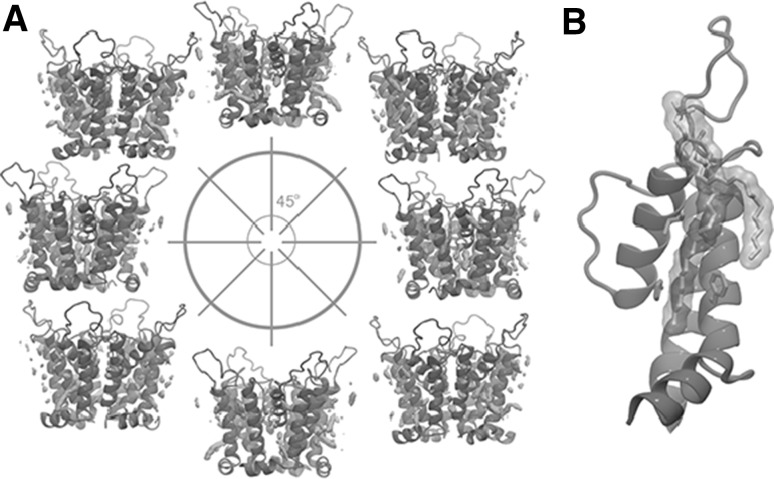
References
-
- Abraham MJ, Murtola T, Schulz R, Páll S, Smith JC, Hess B, Lindahl E. Gromacs: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX. 2015;12:19–25. doi: 10.1016/j.softx.2015.06.001. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
