

Exosome-Mediated Benefits of Cell Therapy in Mouse and Human Models of Duchenne Muscular Dystrophy
- PMID: 29478899
- PMCID: PMC5918344
- DOI: 10.1016/j.stemcr.2018.01.023
Exosome-Mediated Benefits of Cell Therapy in Mouse and Human Models of Duchenne Muscular Dystrophy
Erratum in
-
Exosome-Mediated Benefits of Cell Therapy in Mouse and Human Models of Duchenne Muscular Dystrophy.Stem Cell Reports. 2026 Mar 13:102889. doi: 10.1016/j.stemcr.2026.102889. Online ahead of print. Stem Cell Reports. 2026. PMID: 41831444 No abstract available.
Abstract
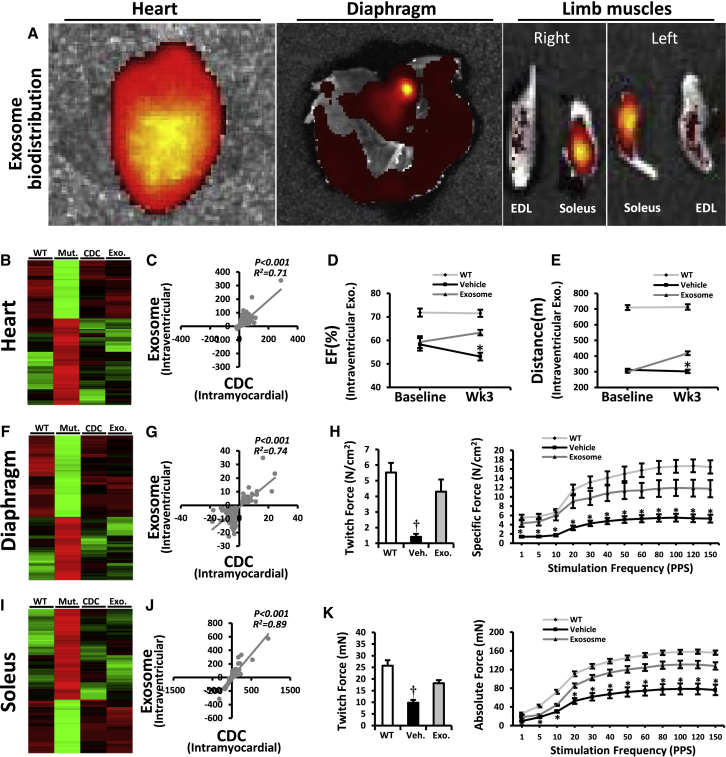
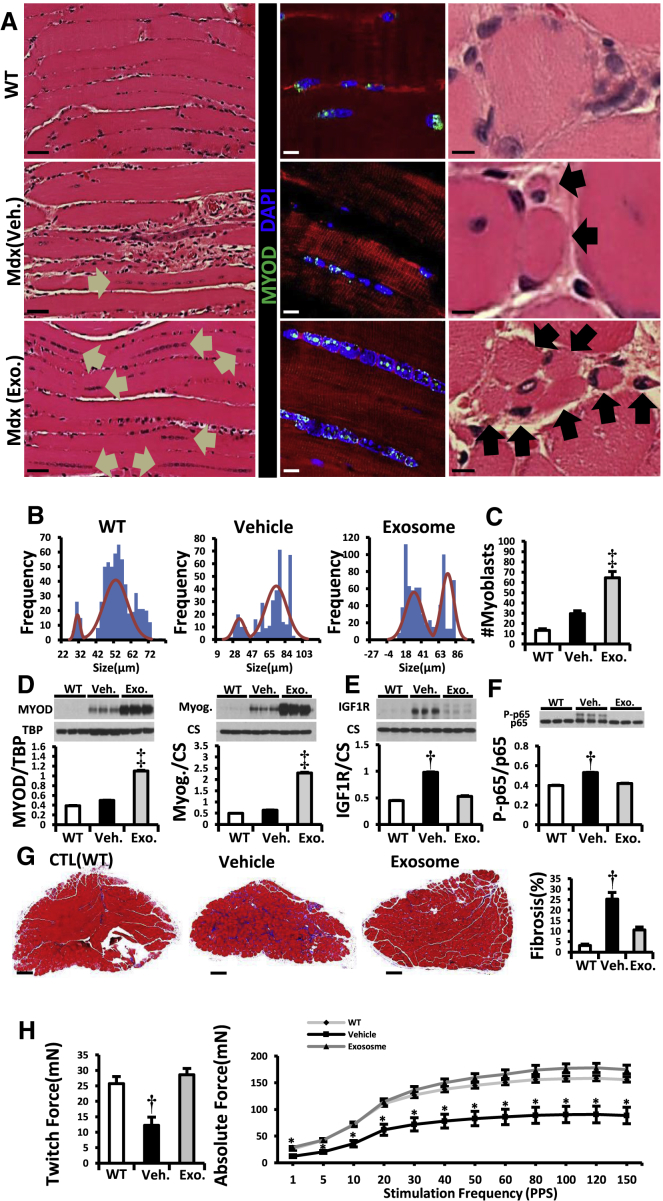
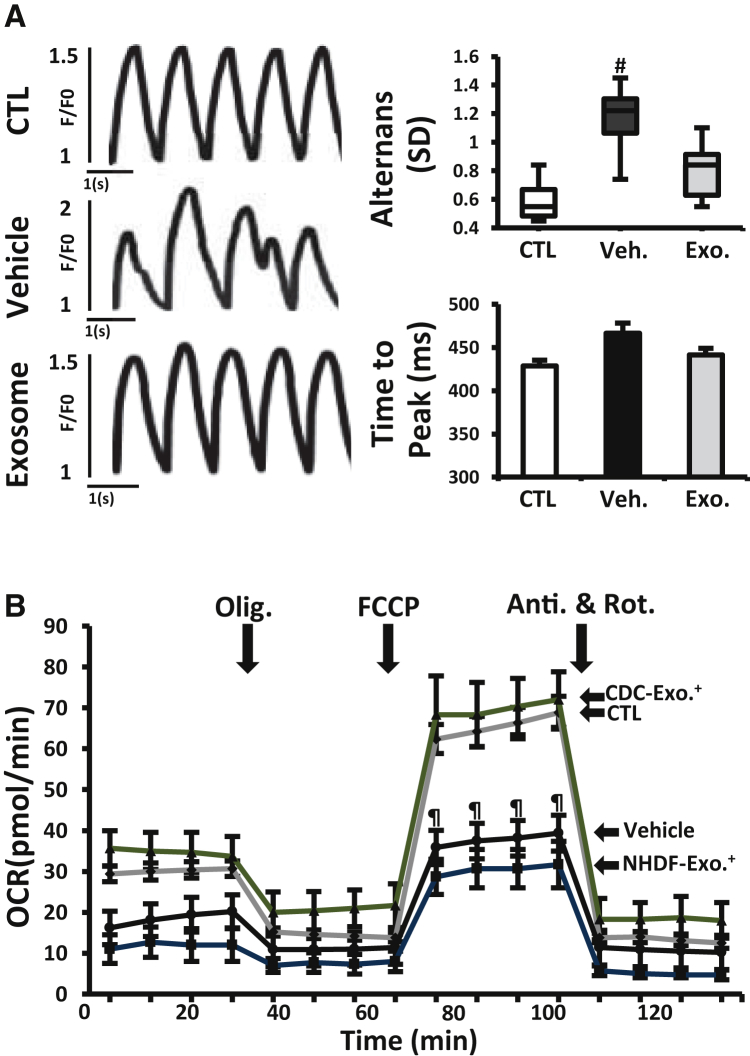
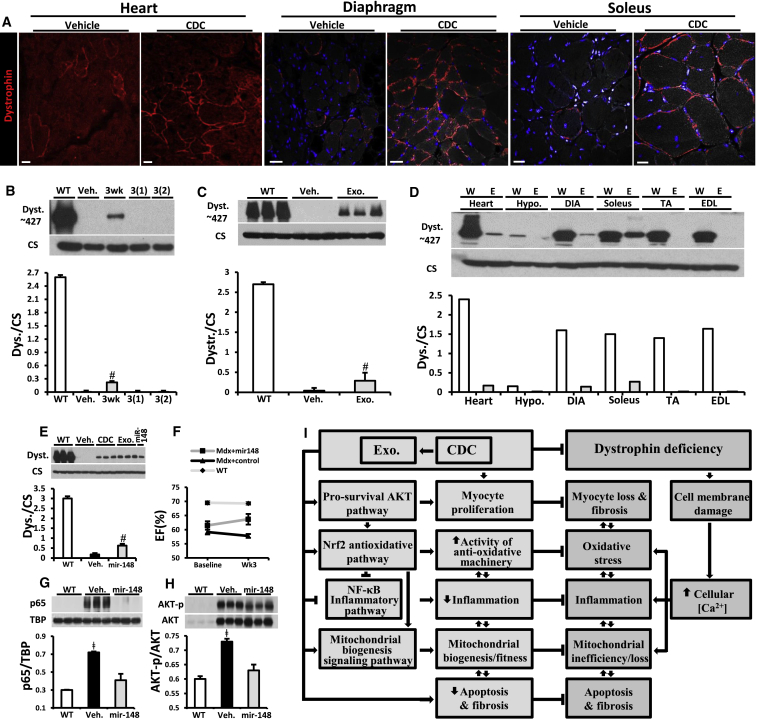
Genetic deficiency of dystrophin leads to disability and premature death in Duchenne muscular dystrophy (DMD), affecting the heart as well as skeletal muscle. Here, we report that clinical-stage cardiac progenitor cells, known as cardiosphere-derived cells (CDCs), improve cardiac and skeletal myopathy in the mdx mouse model of DMD. Injection of CDCs into the hearts of mdx mice augments cardiac function, ambulatory capacity, and survival. Exosomes secreted by human CDCs reproduce the benefits of CDCs in mdx mice and in human induced pluripotent stem cell-derived Duchenne cardiomyocytes. Surprisingly, CDCs and their exosomes also transiently restored partial expression of full-length dystrophin in mdx mice. The findings further motivate the testing of CDCs in Duchenne patients, while identifying exosomes as next-generation therapeutic candidates.
Keywords: cardiomyopathy; cardiosphere-derived cells; dystrophin; exosomes; microRNA; muscular dystrophy.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Aminzadeh M.A., Tobin R., Smith R., Marban L., Marban E. Heart-derived cell therapy for Duchenne cardiomyopathy: cardiosphere-derived cells and their exosomes improve function, restore mitochondrial integrity and reverse degenerative changes in the hearts of Mdx mice. Circ. Res. 2014;115:e90.
-
- Aminzadeh M.A., Durvasula P., Tobin R., Guan X., Andres A., Taylor D., Ibrahim A., Sun B., Torrente A., Goldhaber J. Exosome-mediated reversal of Duchenne cardiomyopathy. Circulation. 2015;132:A16015.
-
- Ascheim D., Jefferies J.L. 2016. A randomized, open-label study of the safety and efficacy of multi-vessel intracoronary delivery of allogeneic cardiosphere-derived cells in patients with cardiomyopathy secondary to Duchenne muscular dystrophy.https://clinicaltrials.gov/ct2/show/NCT02485938
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
