

Stretching the limits: from homeostasis to stem cell plasticity in wound healing and cancer
- PMID: 29479084
- PMCID: PMC6301069
- DOI: 10.1038/nrg.2018.9
Stretching the limits: from homeostasis to stem cell plasticity in wound healing and cancer
Abstract
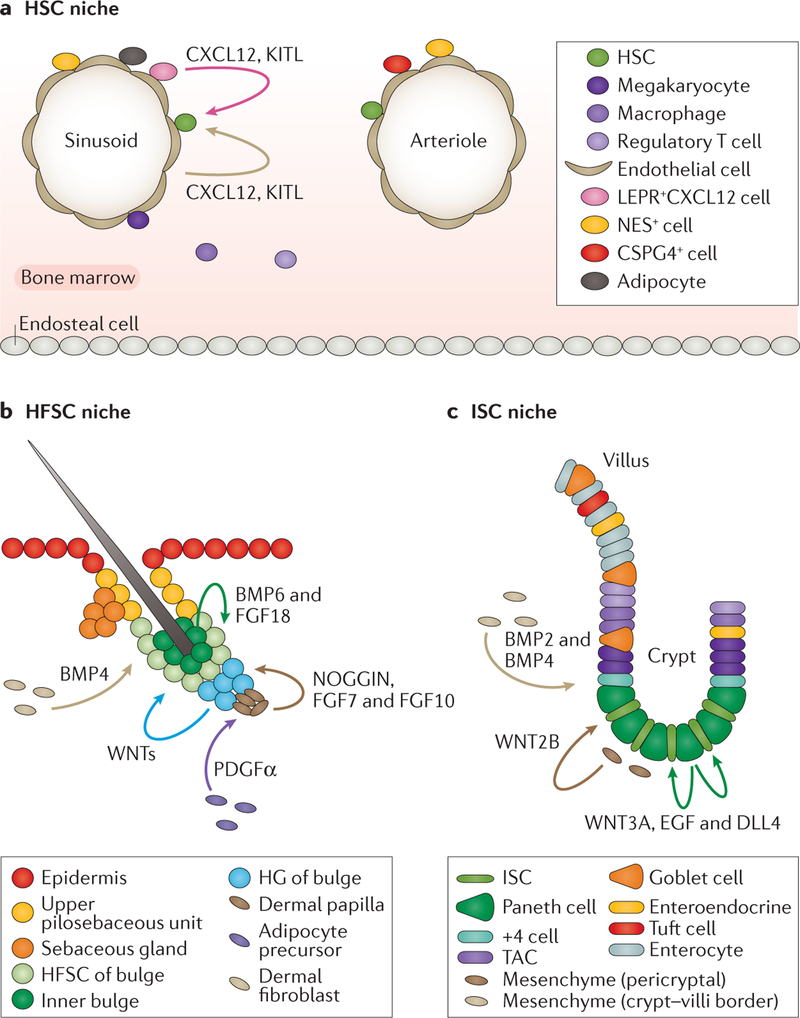
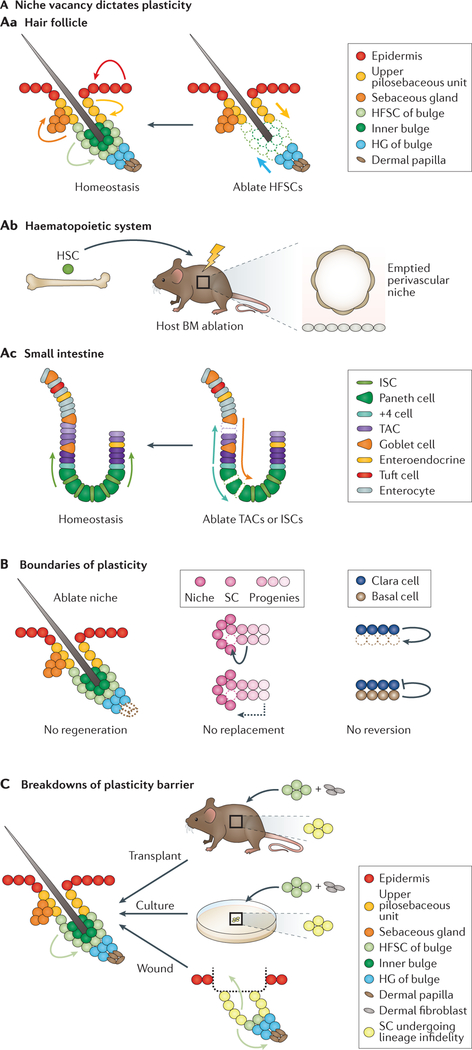
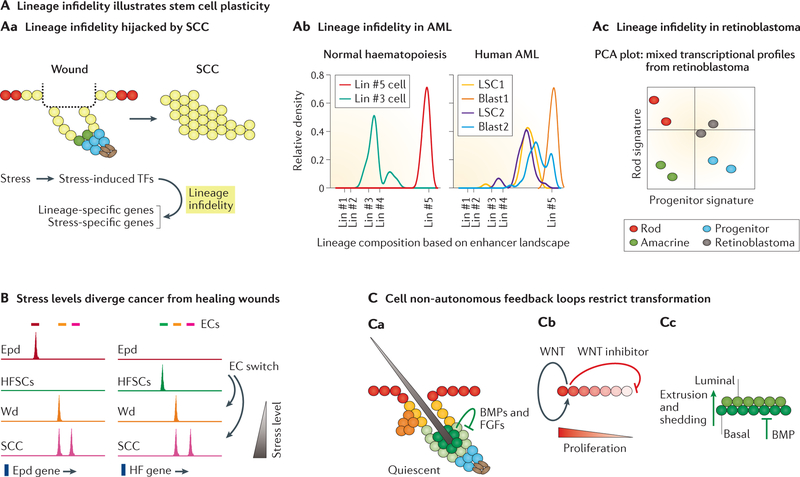
Stem cells (SCs) govern tissue homeostasis and wound repair. They reside within niches, the special microenvironments within tissues that control SC lineage outputs. Upon injury or stress, new signals emanating from damaged tissue can divert nearby cells into adopting behaviours that are not part of their homeostatic repertoire. This behaviour, known as SC plasticity, typically resolves as wounds heal. However, in cancer, it can endure. Recent studies have yielded insights into the orchestrators of maintenance and lineage commitment for SCs belonging to three mammalian tissues: the haematopoietic system, the skin epithelium and the intestinal epithelium. We delineate the multifactorial determinants and general principles underlying the remarkable facets of SC plasticity, which lend promise for regenerative medicine and cancer therapeutics.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
-
Schofield R The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells 4, 7–25 (1978).
In this paper, the SC niche concept is coined and formally elaborated upon.
-
-
-
Till JE & McCulloch EA A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiat. Res 14, 213–222 (1961).
This article presents the discovery of spleen colony-forming units, clonogenic bone marrow precursors that give rise to macroscopic spleen colonies after transplantation into irradiated recipient mice, a work directly leading to the definition of a SC.
-
-
- Dick JE, Magli MC, Huszar D, Phillips RA & Bernstein A Introduction of a selectable gene into primitive stem cells capable of long-term reconstitution of the hemopoietic system of W/Wv mice. Cell 42, 71–79 (1985). - PubMed
-
- Lemischka IR, Raulet DH & Mulligan RC Developmental potential and dynamic behavior of hematopoietic stem cells. Cell 45, 917–927 (1986). - PubMed
-
- Muller-Sieburg CE, Whitlock CA & Weissman IL Isolation of two early B lymphocyte progenitors from mouse marrow: a committed pre-pre-B cell and a clonogenic Thy-1-lo hematopoietic stem cell. Cell 44, 653–662 (1986). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
