

Evaluating Different Virulence Traits of Klebsiella pneumoniae Using Dictyostelium discoideum and Zebrafish Larvae as Host Models
- PMID: 29479519
- PMCID: PMC5811510
- DOI: 10.3389/fcimb.2018.00030
Evaluating Different Virulence Traits of Klebsiella pneumoniae Using Dictyostelium discoideum and Zebrafish Larvae as Host Models
Abstract
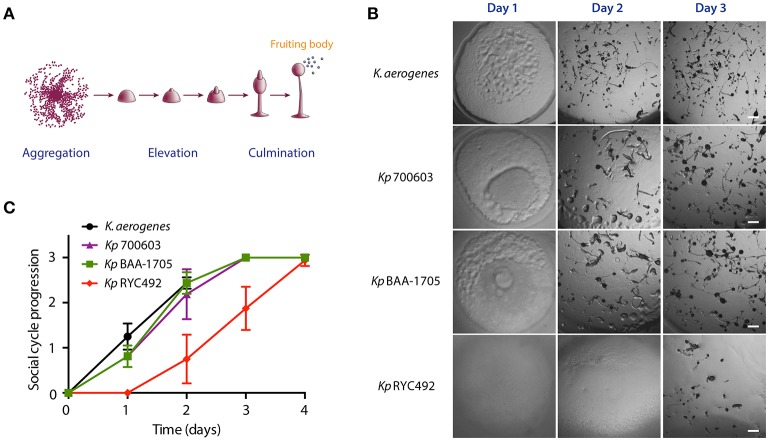
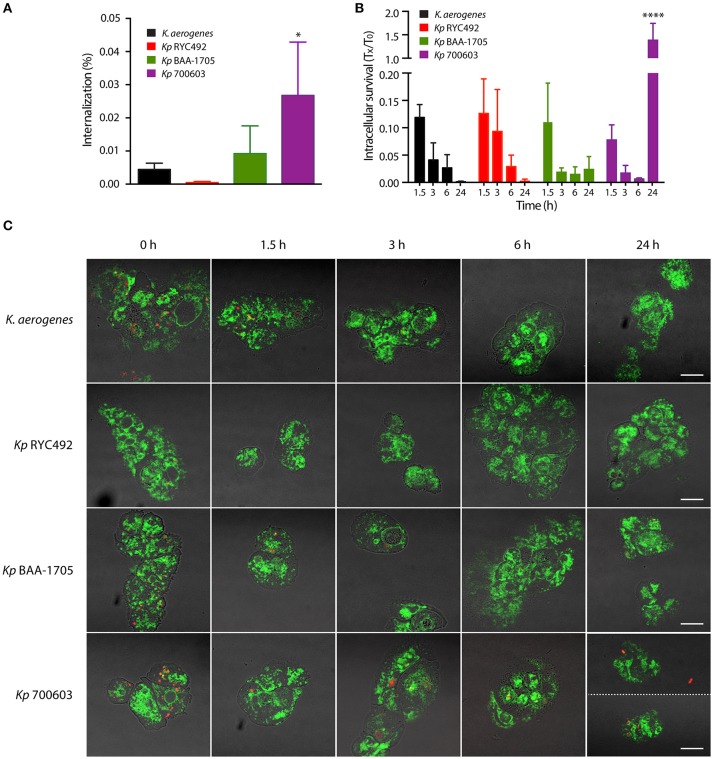
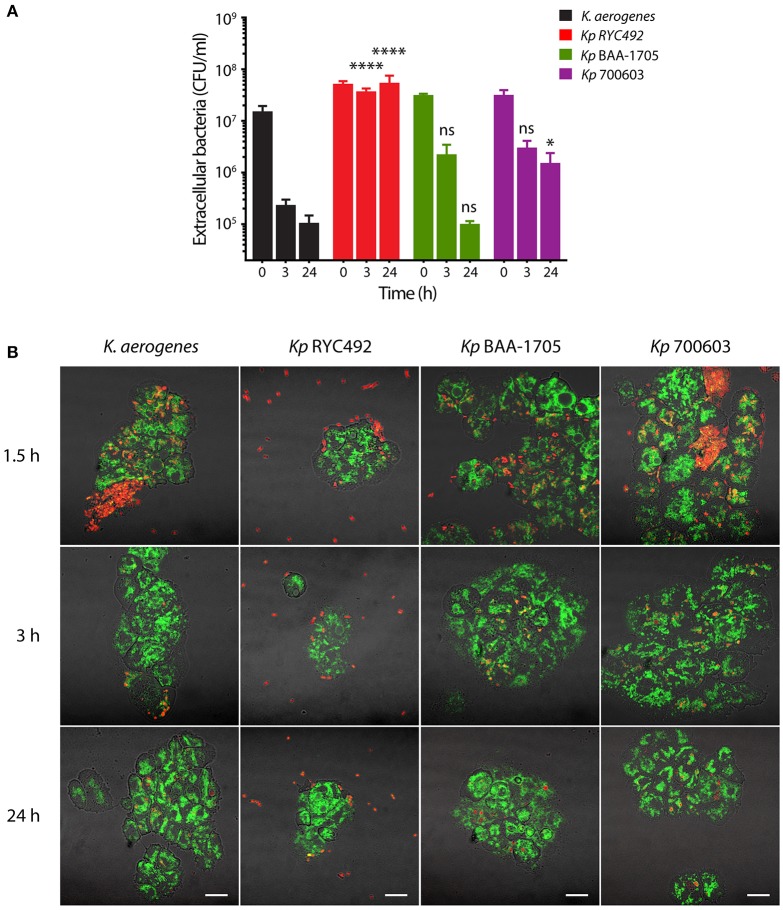
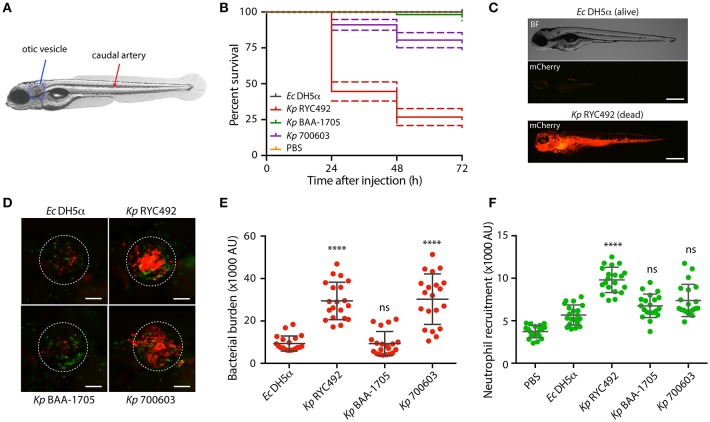
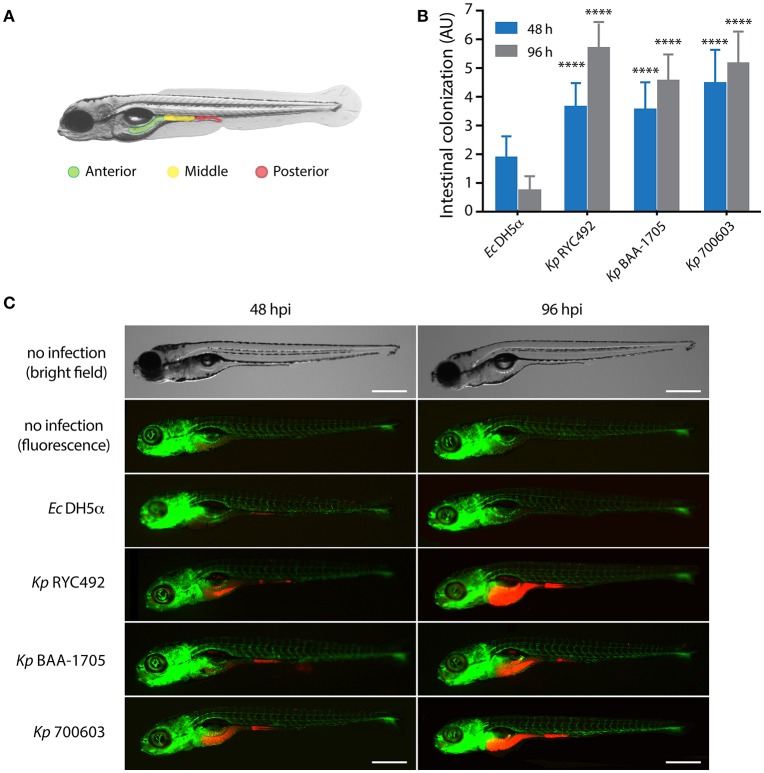
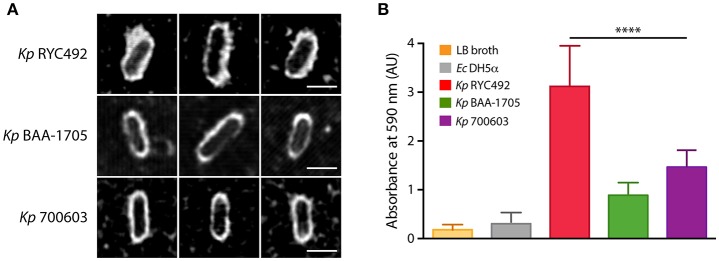
Multiresistant and invasive hypervirulent Klebsiella pneumoniae strains have become one of the most urgent bacterial pathogen threats. Recent analyses revealed a high genomic plasticity of this species, harboring a variety of mobile genetic elements associated with virulent strains, encoding proteins of unknown function whose possible role in pathogenesis have not been addressed. K. pneumoniae virulence has been studied mainly in animal models such as mice and pigs, however, practical, financial, ethical and methodological issues limit the use of mammal hosts. Consequently, the development of simple and cost-effective experimental approaches with alternative host models is needed. In this work we described the use of both, the social amoeba and professional phagocyte Dictyostelium discoideum and the fish Danio rerio (zebrafish) as surrogate host models to study K. pneumoniae virulence. We compared three K. pneumoniae clinical isolates evaluating their resistance to phagocytosis, intracellular survival, lethality, intestinal colonization, and innate immune cells recruitment. Optical transparency of both host models permitted studying the infective process in vivo, following the Klebsiella-host interactions through live-cell imaging. We demonstrated that K. pneumoniae RYC492, but not the multiresistant strains 700603 and BAA-1705, is virulent to both host models and elicits a strong immune response. Moreover, this strain showed a high resistance to phagocytosis by D. discoideum, an increased ability to form biofilms and a more prominent and irregular capsule. Besides, the strain 700603 showed the unique ability to replicate inside amoeba cells. Genomic comparison of the K. pneumoniae strains showed that the RYC492 strain has a higher overall content of virulence factors although no specific genes could be linked to its phagocytosis resistance, nor to the intracellular survival observed for the 700603 strain. Our results indicate that both zebrafish and D. discoideum are advantageous host models to study different traits of K. pneumoniae that are associated with virulence.
Keywords: Danio rerio; Dictyostelium discoideum; host-pathogen interactions; hypervirulent Klebsiella pneumoniae; intracellular survival; resistance to phagocytosis.
Figures
Similar articles
-
Enhanced invasion and survival of antibiotic- resistant Klebsiella pneumoniae pathotypes in host cells and strain-specific replication in blood.Front Cell Infect Microbiol. 2025 Feb 14;15:1522573. doi: 10.3389/fcimb.2025.1522573. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40028183 Free PMC article.
-
Capsular polysaccharide enables Klebsiella pneumoniae to evade phagocytosis by blocking host-bacteria interactions.mBio. 2025 Mar 12;16(3):e0383824. doi: 10.1128/mbio.03838-24. Epub 2025 Feb 14. mBio. 2025. PMID: 39950808 Free PMC article.
-
The Klebsiella pneumoniae YfgL (BamB) lipoprotein contributes to outer membrane protein biogenesis, type-1 fimbriae expression, anti-phagocytosis, and in vivo virulence.Virulence. 2016 Jul 3;7(5):587-601. doi: 10.1080/21505594.2016.1171435. Epub 2016 Mar 30. Virulence. 2016. PMID: 27029012 Free PMC article.
-
Antimicrobial Resistance of Hypervirulent Klebsiella pneumoniae: Epidemiology, Hypervirulence-Associated Determinants, and Resistance Mechanisms.Front Cell Infect Microbiol. 2017 Nov 21;7:483. doi: 10.3389/fcimb.2017.00483. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 29209595 Free PMC article. Review.
-
Finding Order in the Chaos: Outstanding Questions in Klebsiella pneumoniae Pathogenesis.Infect Immun. 2021 Mar 17;89(4):e00693-20. doi: 10.1128/IAI.00693-20. Print 2021 Mar 17. Infect Immun. 2021. PMID: 33558323 Free PMC article. Review.
Cited by
-
Adherence to Human Colon Cells by Multidrug Resistant Enterobacterales Strains Isolated From Solid Organ Transplant Recipients With a Focus on Citrobacter freundii.Front Cell Infect Microbiol. 2020 Sep 16;10:447. doi: 10.3389/fcimb.2020.00447. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33042855 Free PMC article.
-
The Ferric uptake regulator (Fur) and iron availability control the production and maturation of the antibacterial peptide microcin E492.PLoS One. 2018 Aug 2;13(8):e0200835. doi: 10.1371/journal.pone.0200835. eCollection 2018. PLoS One. 2018. PMID: 30071030 Free PMC article.
-
Ultrashort Cationic Lipopeptides-Effect of N-Terminal Amino Acid and Fatty Acid Type on Antimicrobial Activity and Hemolysis.Molecules. 2020 Jan 8;25(2):257. doi: 10.3390/molecules25020257. Molecules. 2020. PMID: 31936341 Free PMC article.
-
Baicalein Resensitizes Multidrug-Resistant Gram-Negative Pathogens to Doxycycline.Microbiol Spectr. 2023 Jun 15;11(3):e0470222. doi: 10.1128/spectrum.04702-22. Epub 2023 Apr 18. Microbiol Spectr. 2023. PMID: 37070985 Free PMC article.
-
Contribution of the Twin-Arginine Translocation System to the Intracellular Survival of Salmonella Typhimurium in Dictyostelium discoideum.Front Microbiol. 2018 Dec 6;9:3001. doi: 10.3389/fmicb.2018.03001. eCollection 2018. Front Microbiol. 2018. PMID: 30574134 Free PMC article.
References
-
- Balestrino D., Ghigo J. M., Charbonnel N., Haagensen J. A., Forestier C. (2008). The characterization of functions involved in the establishment and maturation of Klebsiella pneumoniae in vitro biofilm reveals dual roles for surface exopolysaccharides. Environ. Microbiol. 10, 685–701. 10.1111/j.1462-2920.2007.01491.x - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials