

Timing of host feeding drives rhythms in parasite replication
- PMID: 29481559
- PMCID: PMC5843352
- DOI: 10.1371/journal.ppat.1006900
Timing of host feeding drives rhythms in parasite replication
Abstract
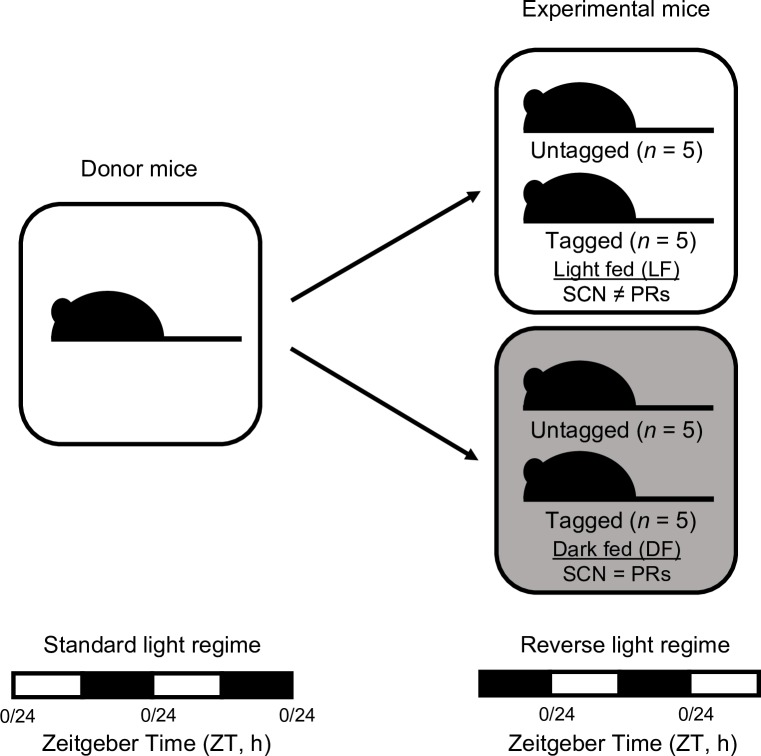
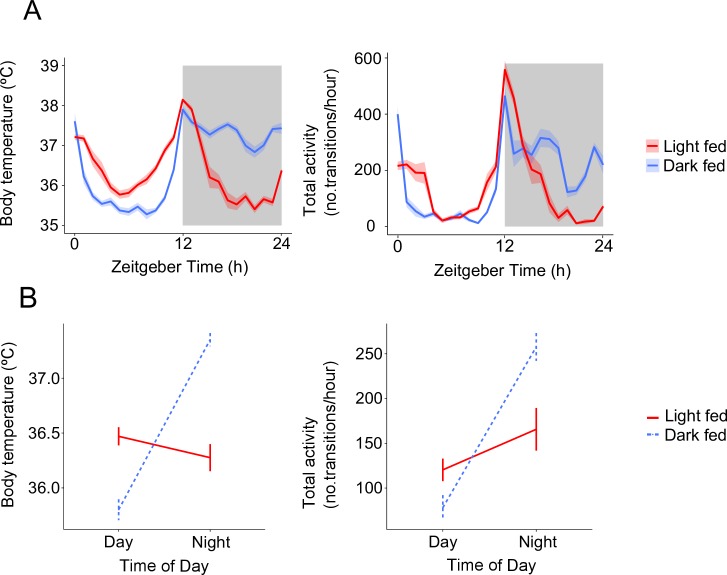
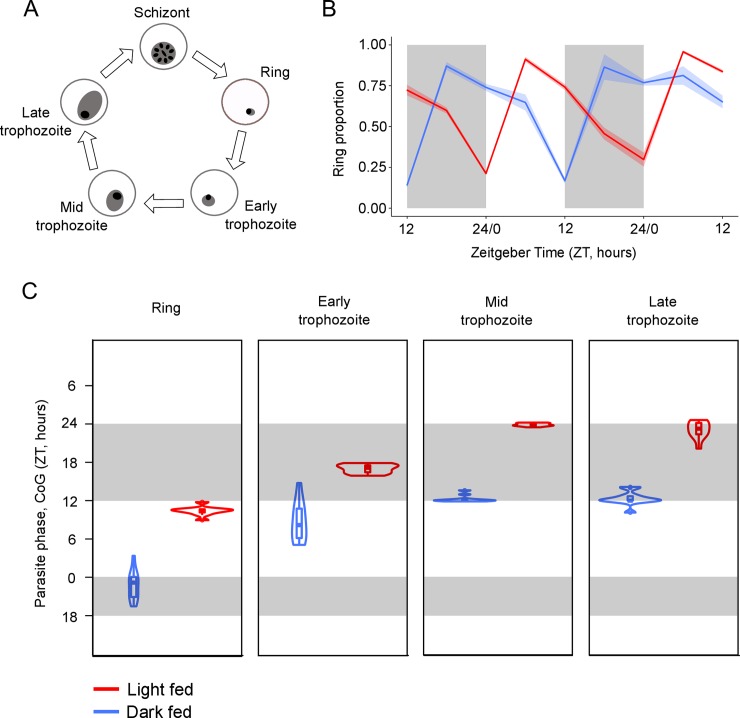
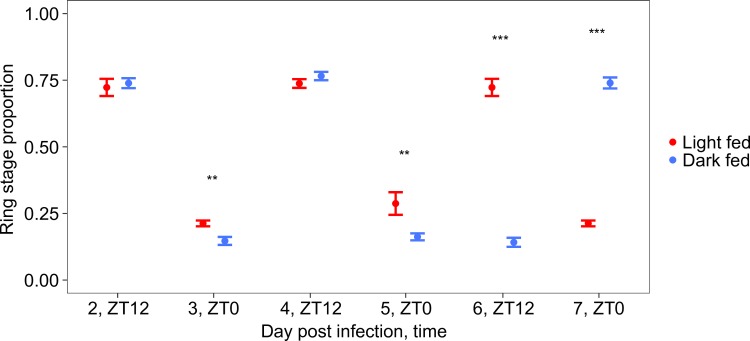
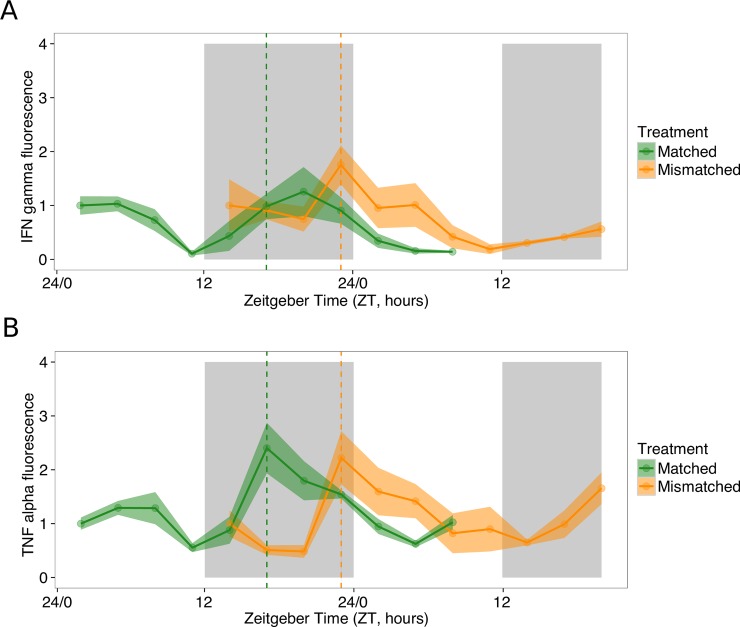
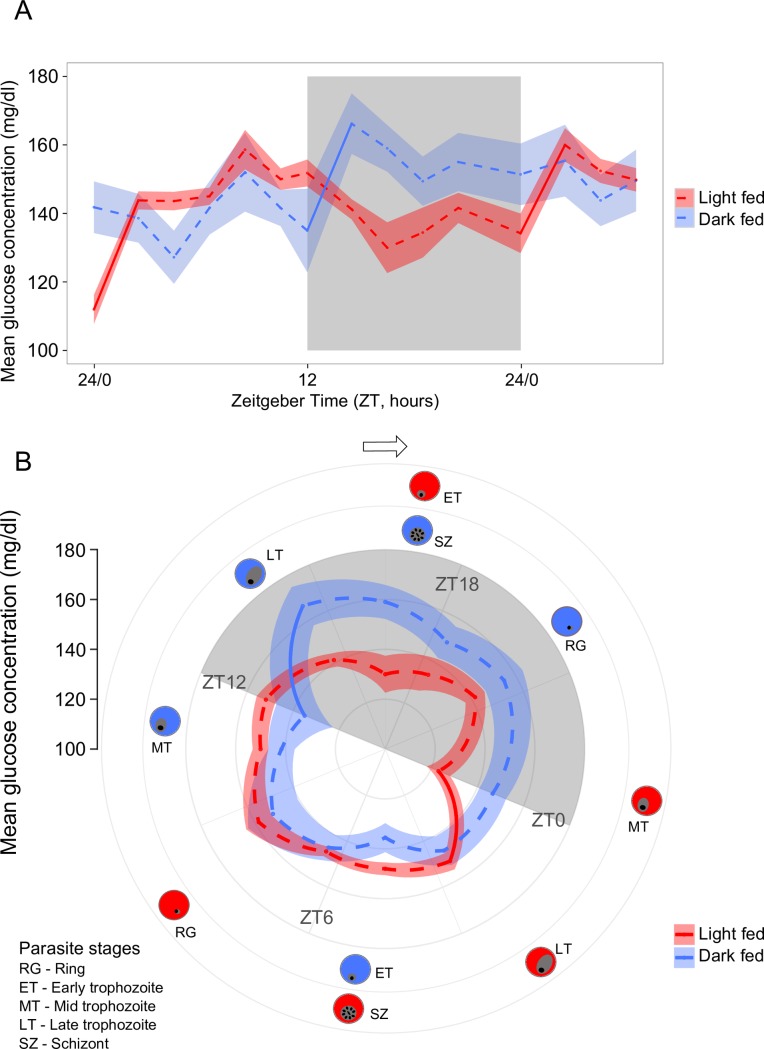
Circadian rhythms enable organisms to synchronise the processes underpinning survival and reproduction to anticipate daily changes in the external environment. Recent work shows that daily (circadian) rhythms also enable parasites to maximise fitness in the context of ecological interactions with their hosts. Because parasite rhythms matter for their fitness, understanding how they are regulated could lead to innovative ways to reduce the severity and spread of diseases. Here, we examine how host circadian rhythms influence rhythms in the asexual replication of malaria parasites. Asexual replication is responsible for the severity of malaria and fuels transmission of the disease, yet, how parasite rhythms are driven remains a mystery. We perturbed feeding rhythms of hosts by 12 hours (i.e. diurnal feeding in nocturnal mice) to desynchronise the host's peripheral oscillators from the central, light-entrained oscillator in the brain and their rhythmic outputs. We demonstrate that the rhythms of rodent malaria parasites in day-fed hosts become inverted relative to the rhythms of parasites in night-fed hosts. Our results reveal that the host's peripheral rhythms (associated with the timing of feeding and metabolism), but not rhythms driven by the central, light-entrained circadian oscillator in the brain, determine the timing (phase) of parasite rhythms. Further investigation reveals that parasite rhythms correlate closely with blood glucose rhythms. In addition, we show that parasite rhythms resynchronise to the altered host feeding rhythms when food availability is shifted, which is not mediated through rhythms in the host immune system. Our observations suggest that parasites actively control their developmental rhythms. Finally, counter to expectation, the severity of disease symptoms expressed by hosts was not affected by desynchronisation of their central and peripheral rhythms. Our study at the intersection of disease ecology and chronobiology opens up a new arena for studying host-parasite-vector coevolution and has broad implications for applied bioscience.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hevia MA, Canessa P, Müller-Esparza H, Larrondo LF. A circadian oscillator in the fungus Botrytis cinerea regulates virulence when infecting Arabidopsis thaliana. PNAS. 2015;112: 8744–8749. doi: 10.1073/pnas.1508432112 - DOI - PMC - PubMed
-
- N’Goran E, Brémond P, Sellin E, Sellin B, Théron A. Intraspecific diversity of Schistosoma haematobium in West Africa: chronobiology of cercarial emergence. Acta Trop. 1997;66: 35–44. - PubMed
-
- Dolnik OV, Metzger BJ, Loonen MJJE. Keeping the clock set under the midnight sun: diurnal periodicity and synchrony of avian Isospora parasites cycle in the High Arctic. Parasitol. 2011;138: 1077–1081. - PubMed
-
- Hawking F. The 24-hour periodicity of microfilariae: biological mechanisms responsible for its production and control. Proc Roy Soc B. 1967;169: 59–76.
-
- Hawking F. Circadian rhythms of Trypanosoma congolense in laboratory rodents. Trans R Soc Trop Med Hyg. 1978;72: 592–595. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
