

Microbial co-infection alters macrophage polarization, phagosomal escape, and microbial killing
- PMID: 29482417
- PMCID: PMC6852389
- DOI: 10.1177/1753425918760180
Microbial co-infection alters macrophage polarization, phagosomal escape, and microbial killing
Abstract
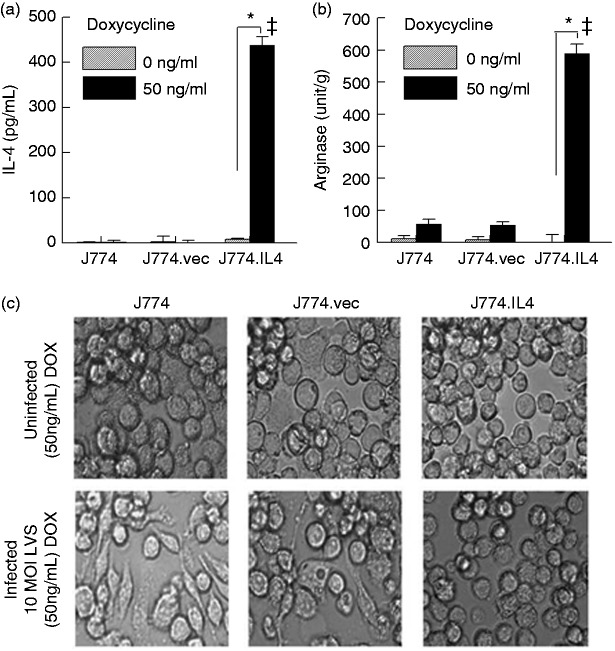
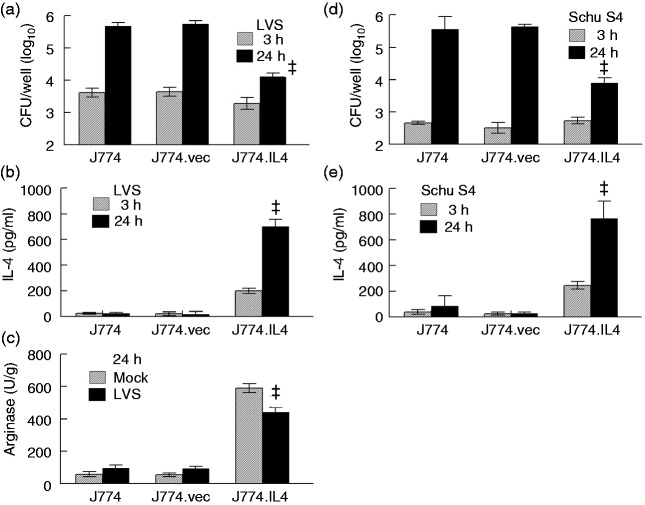
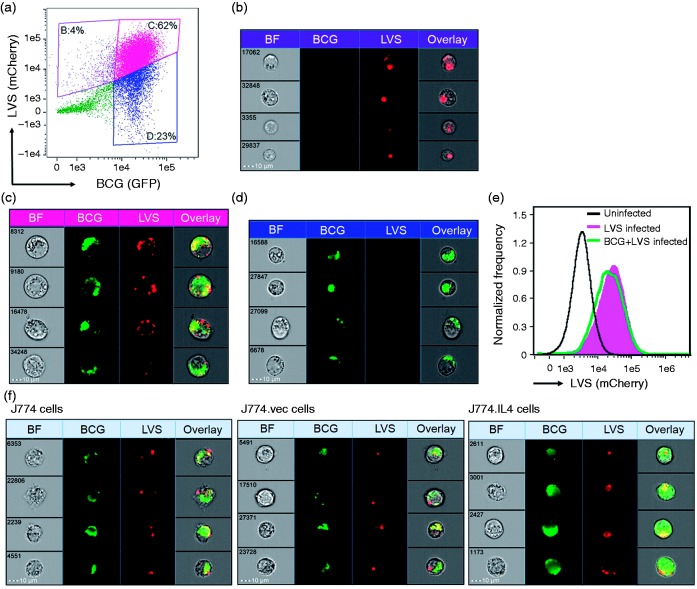
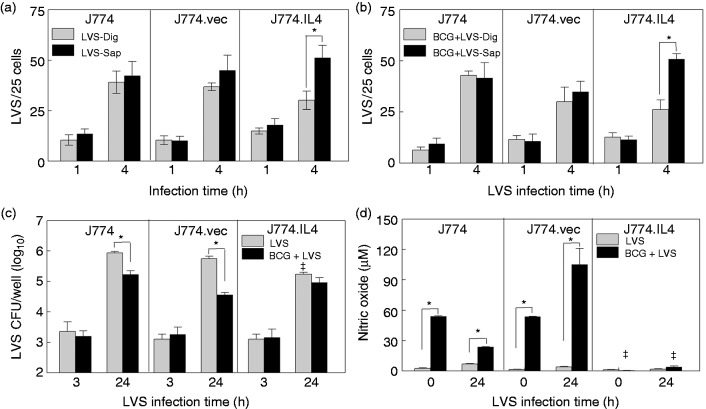
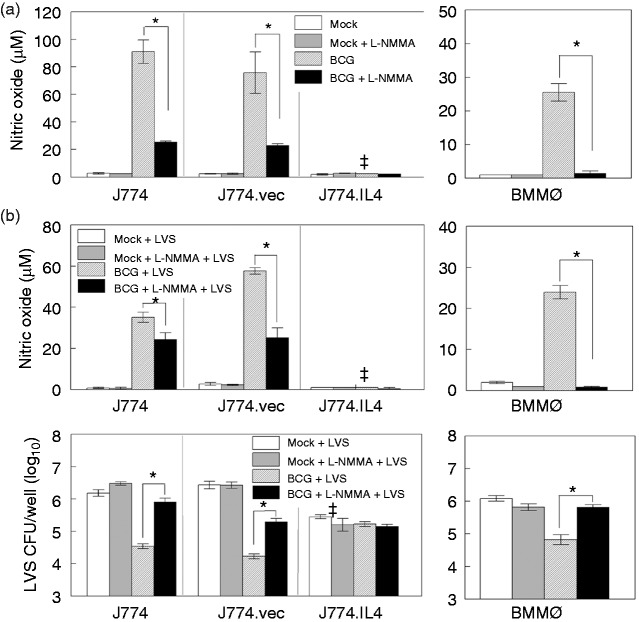
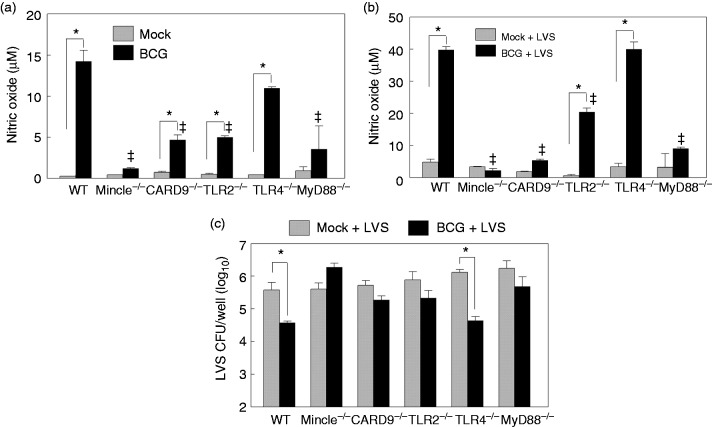
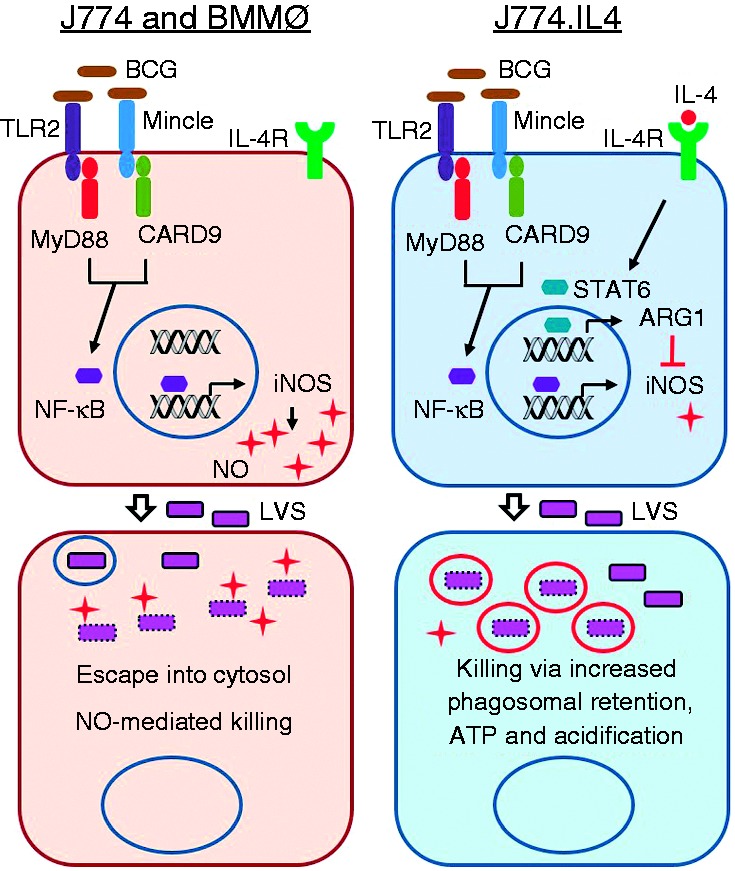
Macrophages are important innate immune cells that respond to microbial insults. In response to multi-bacterial infection, the macrophage activation state may change upon exposure to nascent mediators, which results in different bacterial killing mechanism(s). In this study, we utilized two respiratory bacterial pathogens, Mycobacterium bovis (Bacillus Calmette Guẻrin, BCG) and Francisella tularensis live vaccine strain (LVS) with different phagocyte evasion mechanisms, as model microbes to assess the influence of initial bacterial infection on the macrophage response to secondary infection. Non-activated (M0) macrophages or activated M2-polarized cells (J774 cells transfected with the mouse IL-4 gene) were first infected with BCG for 24-48 h, subsequently challenged with LVS, and the results of inhibition of LVS replication in the macrophages was assessed. BCG infection in M0 macrophages activated TLR2-MyD88 and Mincle-CARD9 signaling pathways, stimulating nitric oxide (NO) production and enhanced killing of LVS. BCG infection had little effect on LVS escape from phagosomes into the cytosol in M0 macrophages. In contrast, M2-polarized macrophages exhibited enhanced endosomal acidification, as well as inhibiting LVS replication. Pre-infection with BCG did not induce NO production and thus did not further reduce LVS replication. This study provides a model for studies of the complexity of macrophage activation in response to multi-bacterial infection.
Keywords: BCG; Francisella; IL-4; co-infection; macrophage.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures
References
-
- Arrevillaga G, Gaona J, Sanchez C, et al. Respiratory syncytial virus persistence in macrophages downregulates intercellular adhesion molecule-1 expression and reduces adhesion of non-typeable Haemophilus influenzae. Intervirology 2012; 55: 442–450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
