

Distinct core promoter codes drive transcription initiation at key developmental transitions in a marine chordate
- PMID: 29482522
- PMCID: PMC6389100
- DOI: 10.1186/s12864-018-4504-5
Distinct core promoter codes drive transcription initiation at key developmental transitions in a marine chordate
Abstract
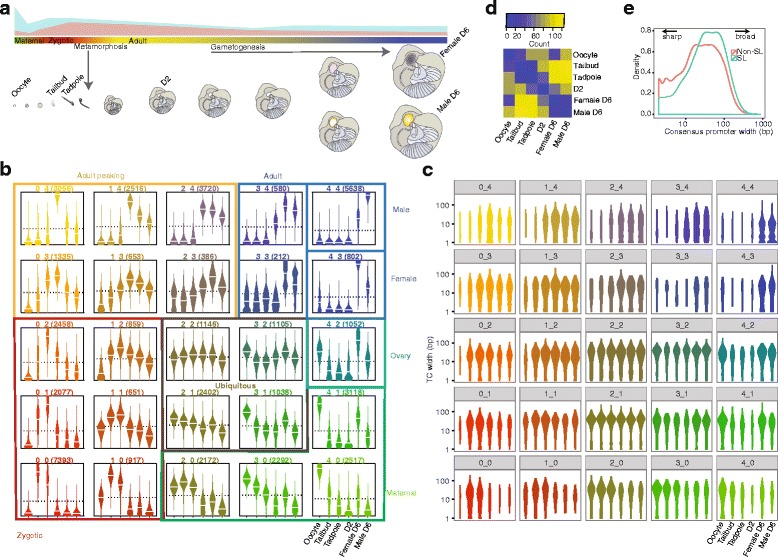
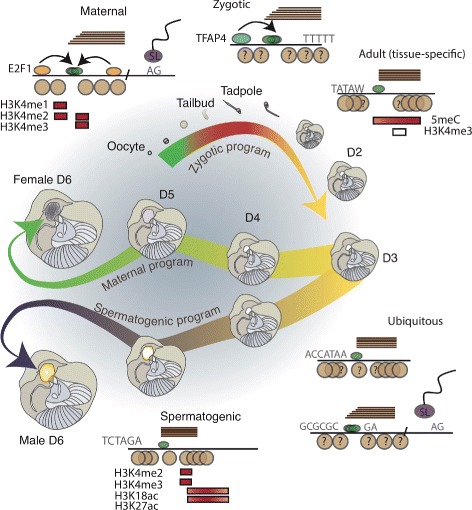
Background: Development is largely driven by transitions between transcriptional programs. The initiation of transcription at appropriate sites in the genome is a key component of this and yet few rules governing selection are known. Here, we used cap analysis of gene expression (CAGE) to generate bp-resolution maps of transcription start sites (TSSs) across the genome of Oikopleura dioica, a member of the closest living relatives to vertebrates.
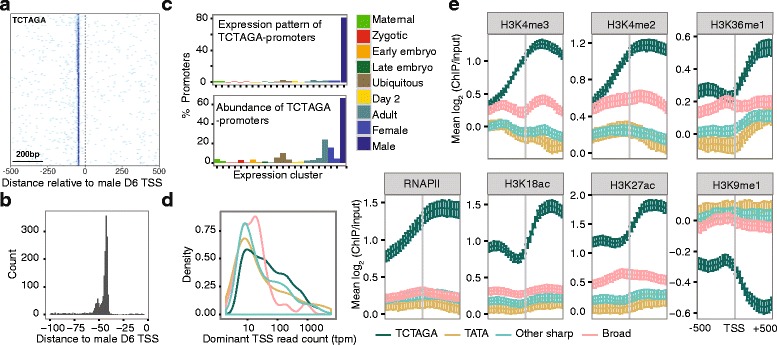
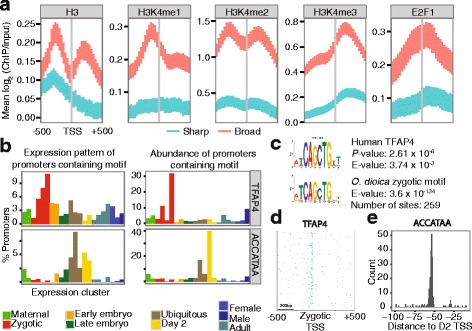
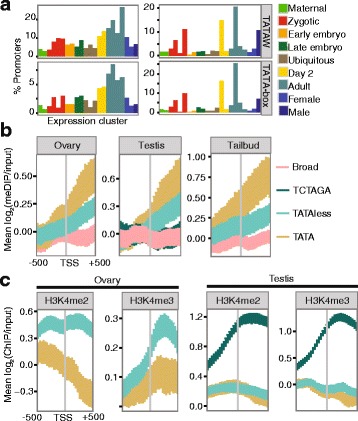
Results: Our TSS maps revealed promoter features in common with vertebrates, as well as striking differences, and uncovered key roles for core promoter elements in the regulation of development. During spermatogenesis there is a genome-wide shift in mode of transcription initiation characterized by a novel core promoter element. This element was associated with > 70% of male-specific transcription, including the use of cryptic internal promoters within operons. In many cases this led to the exclusion of trans-splice sites, revealing a novel mechanism for regulating which mRNAs receive the spliced leader. Binding of the cell cycle regulator, E2F1, is enriched at the TSS of maternal genes in endocycling nurse nuclei. In addition, maternal promoters lack the TATA-like element found in zebrafish and have broad, rather than sharp, architectures with ordered nucleosomes. Promoters of ribosomal protein genes lack the highly conserved TCT initiator. We also report an association between DNA methylation on transcribed gene bodies and the TATA-box.
Conclusions: Our results reveal that distinct functional promoter classes and overlapping promoter codes are present in protochordates like in vertebrates, but show extraordinary lineage-specific innovations. Furthermore, we uncover a genome-wide, developmental stage-specific shift in the mode of TSS selection. Our results provide a rich resource for the study of promoter structure and evolution in Metazoa.
Keywords: Core promoter; DNA methylation; Histone modification; MZT; Oogenesis; Operons; Spermatogenesis; TATA-box; Transcription initiation.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Haberle V, Li N, Hadzhiev Y, Plessy C, Previti C, Nepal C, Gehrig J, Dong X, Akalin A, Suzuki AM, van IJcken WFJ, Armant O, Ferg M, Strähle U, Carninci P, Muller F, Lenhard B. Two independent transcription initiation codes overlap on vertebrate core promoters. Nature. 2014;507(7492):381–5. doi: 10.1038/nature12974. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
