

C. elegans ADARs antagonize silencing of cellular dsRNAs by the antiviral RNAi pathway
- PMID: 29483152
- PMCID: PMC5859968
- DOI: 10.1101/gad.310672.117
C. elegans ADARs antagonize silencing of cellular dsRNAs by the antiviral RNAi pathway
Abstract
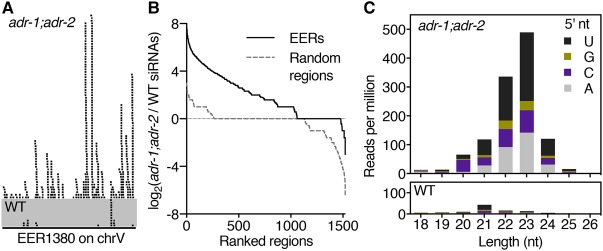
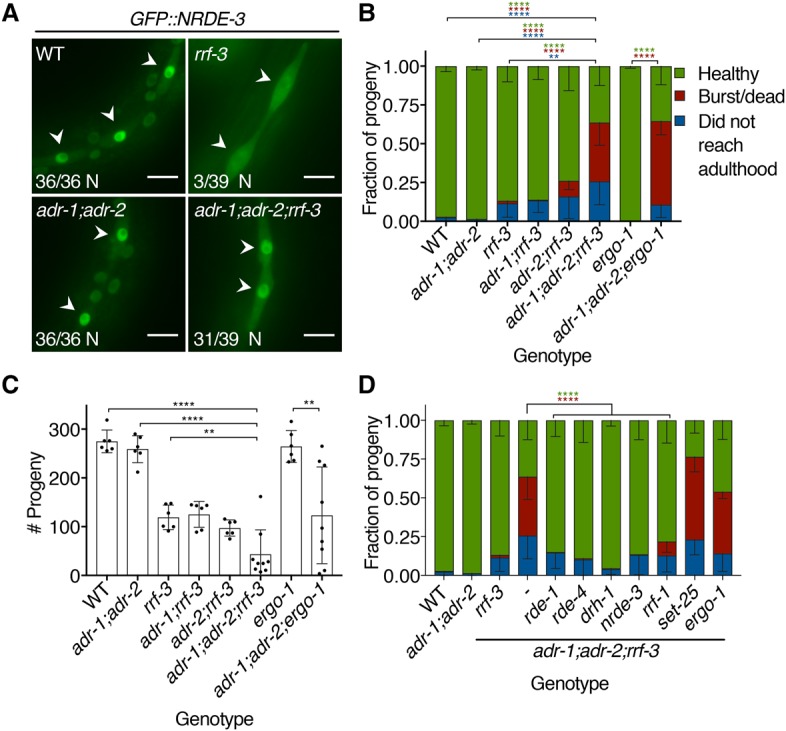
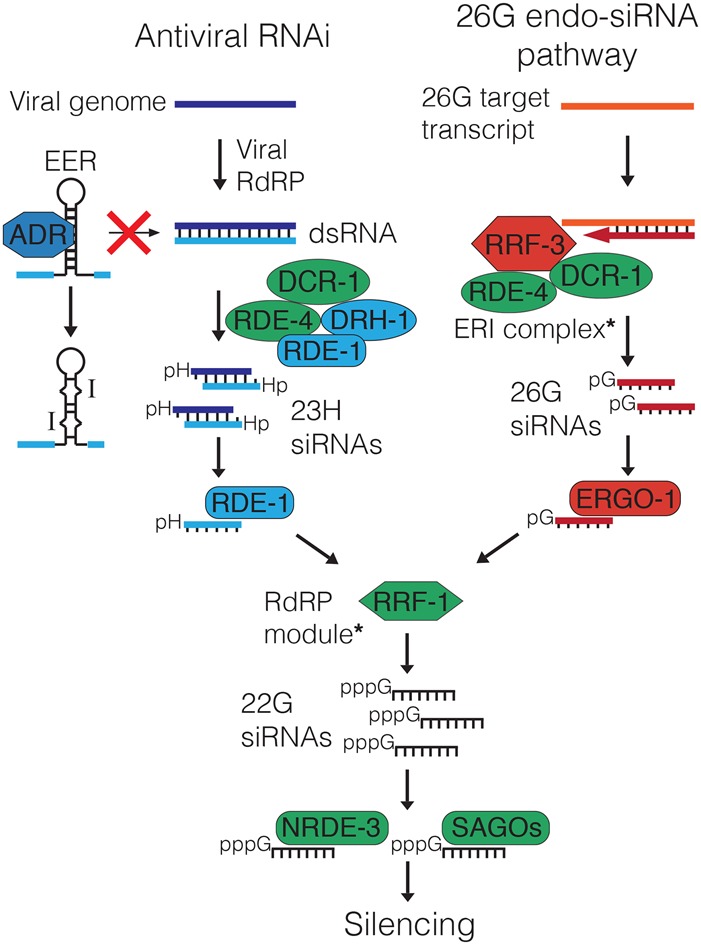
Cellular dsRNAs are edited by adenosine deaminases that act on RNA (ADARs). While editing can alter mRNA-coding potential, most editing occurs in noncoding sequences, the function of which is poorly understood. Using dsRNA immunoprecipitation (dsRIP) and RNA sequencing (RNA-seq), we identified 1523 regions of clustered A-to-I editing, termed editing-enriched regions (EERs), in four stages of Caenorhabditis elegans development, often with highest expression in embryos. Analyses of small RNA-seq data revealed 22- to 23-nucleotide (nt) siRNAs, reminiscent of viral siRNAs, that mapped to EERs and were abundant in adr-1;adr-2 mutant animals. Consistent with roles for these siRNAs in silencing, EER-associated genes (EAGs) were down-regulated in adr-1;adr-2 embryos, and this was dependent on associated EERs and the RNAi factor RDE-4. We observed that ADARs genetically interact with the 26G endogenous siRNA (endo-siRNA) pathway, which likely competes for RNAi components; deletion of factors required for this pathway (rrf-3 or ergo-1) in adr-1;adr-2 mutant strains caused a synthetic phenotype that was rescued by deleting antiviral RNAi factors. Poly(A)+ RNA-seq revealed EAG down-regulation and antiviral gene induction in adr-1;adr-2;rrf-3 embryos, and these expression changes were dependent on rde-1 and rde-4 Our data suggest that ADARs restrict antiviral silencing of cellular dsRNAs.
Keywords: Dicer; RNA editing; deaminase; self–nonself; siRNA.
© 2018 Reich et al.; Published by Cold Spring Harbor Laboratory Press.
Figures



Comment in
-
A rADAR defense against RNAi.Genes Dev. 2018 Feb 1;32(3-4):199-201. doi: 10.1101/gad.313049.118. Genes Dev. 2018. PMID: 29491134 Free PMC article.
References
-
- Billi AC, Fischer SE, Kim JK. 2014. Endogenous RNAi pathways in C. elegans. WormBook (ed. The C. elegans Research Community). WormBook 10.1895/wormbook.1.170.1, http://www.wormbook.org. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials