

Self-interaction of NPM1 modulates multiple mechanisms of liquid-liquid phase separation
- PMID: 29483575
- PMCID: PMC5827731
- DOI: 10.1038/s41467-018-03255-3
Self-interaction of NPM1 modulates multiple mechanisms of liquid-liquid phase separation
Abstract
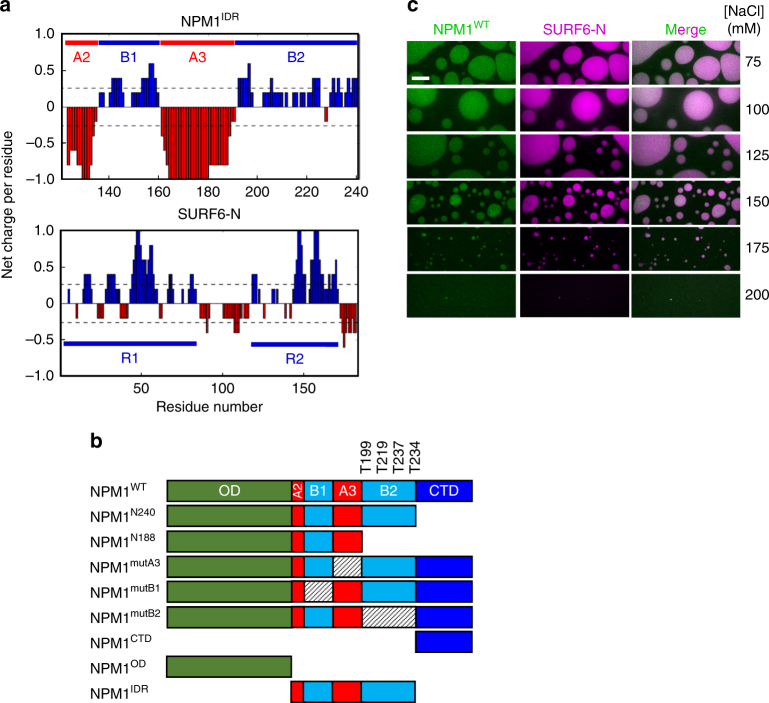
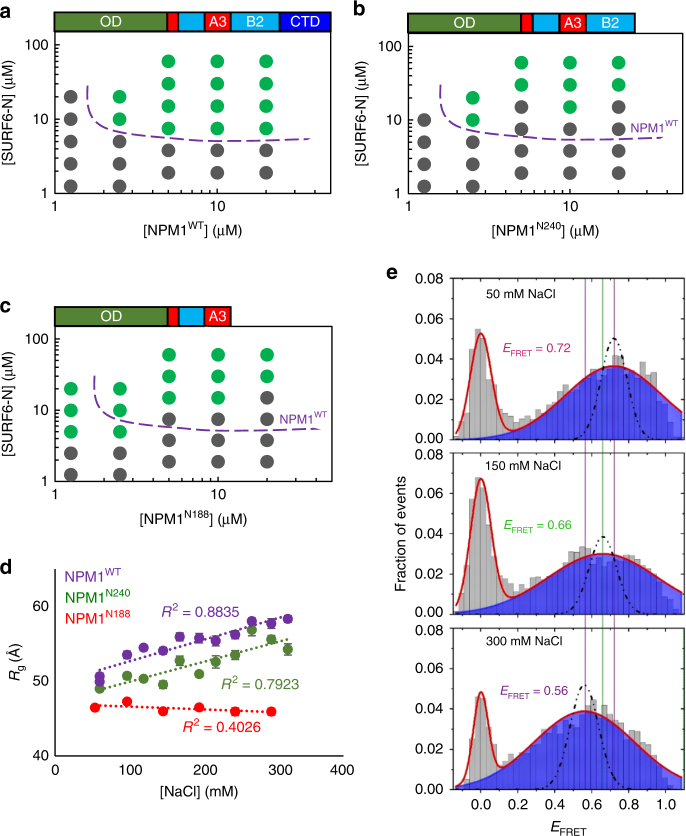
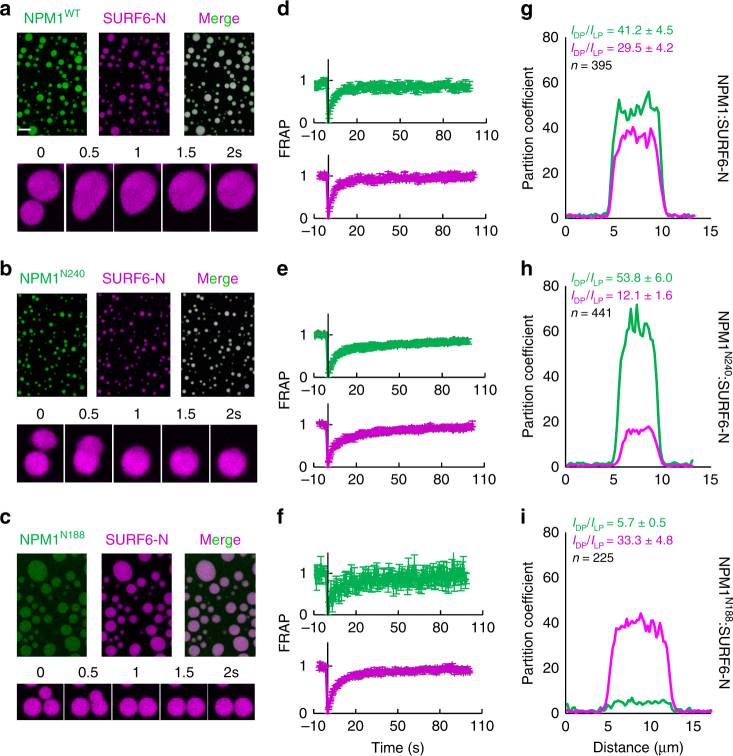
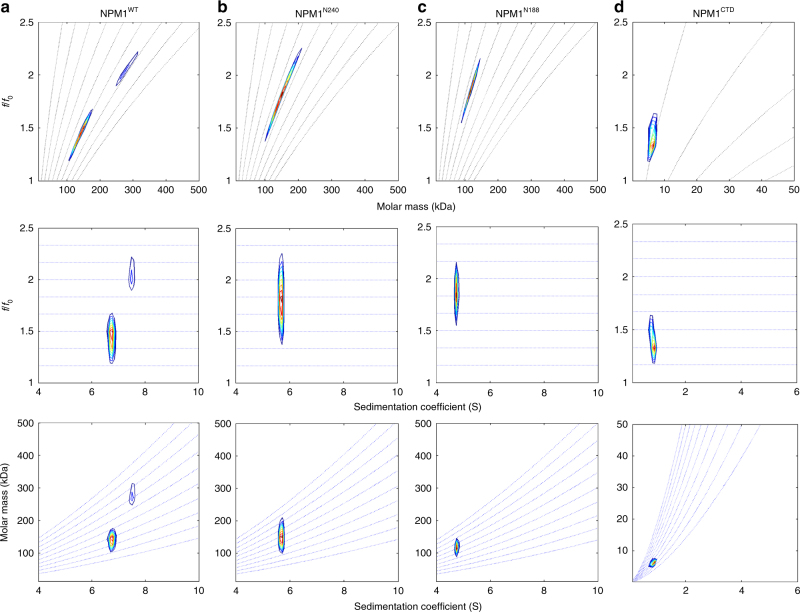
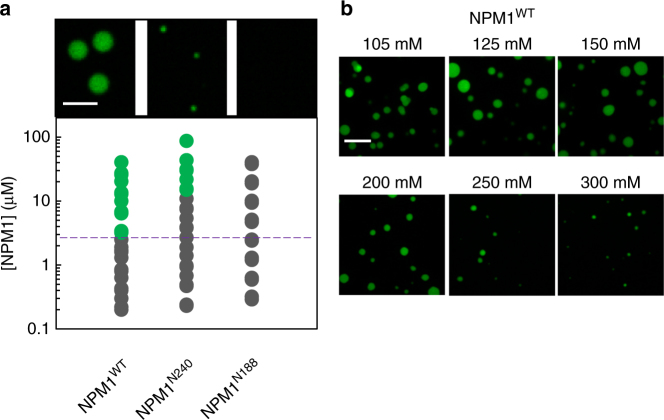
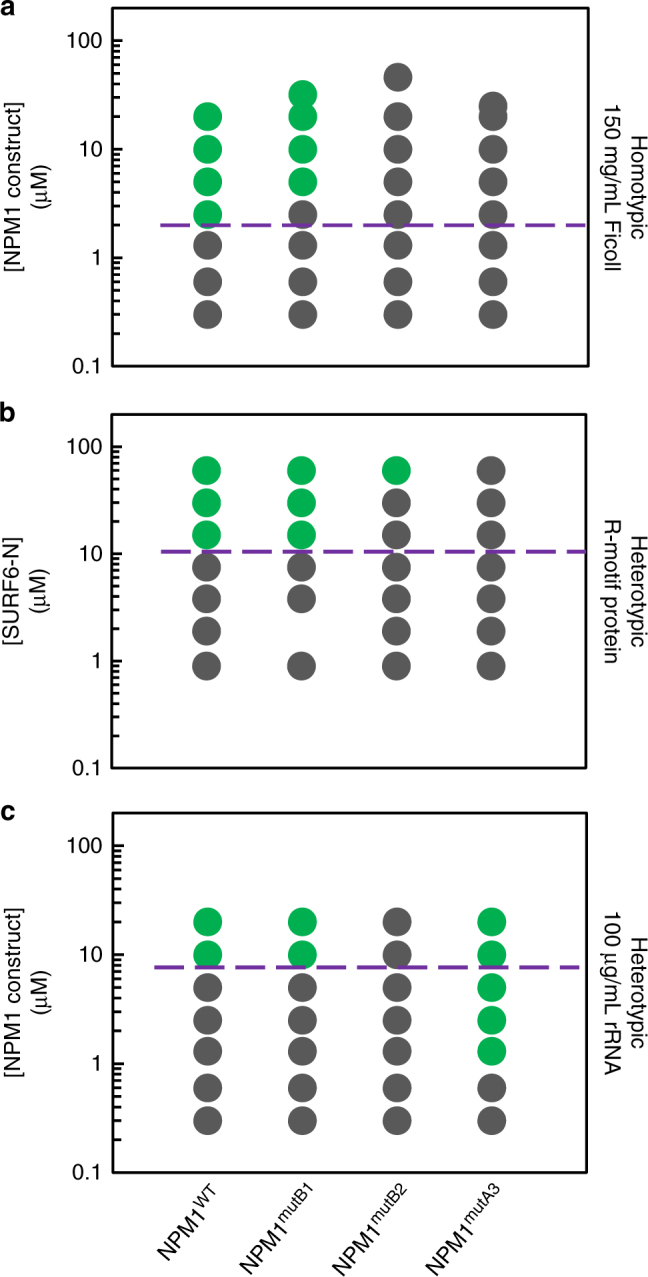
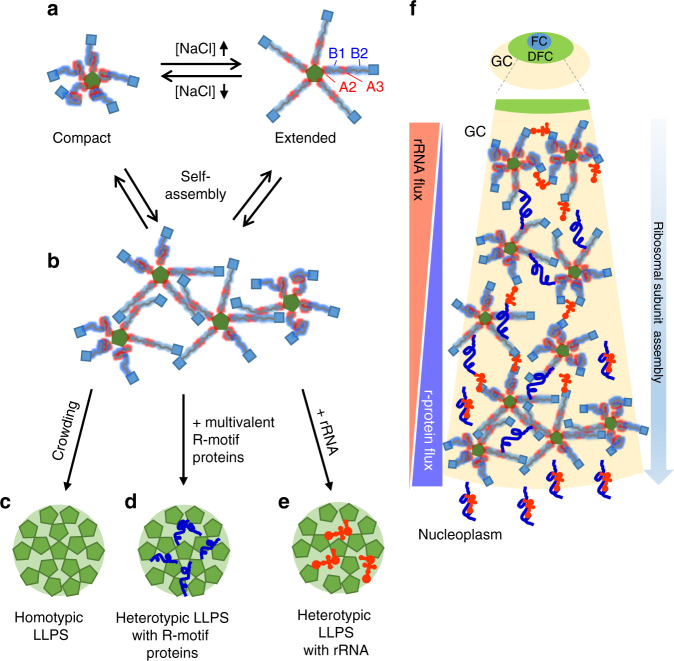
Nucleophosmin (NPM1) is an abundant, oligomeric protein in the granular component of the nucleolus with roles in ribosome biogenesis. Pentameric NPM1 undergoes liquid-liquid phase separation (LLPS) via heterotypic interactions with nucleolar components, including ribosomal RNA (rRNA) and proteins which display multivalent arginine-rich linear motifs (R-motifs), and is integral to the liquid-like nucleolar matrix. Here we show that NPM1 can also undergo LLPS via homotypic interactions between its polyampholytic intrinsically disordered regions, a mechanism that opposes LLPS via heterotypic interactions. Using a combination of biophysical techniques, including confocal microscopy, SAXS, analytical ultracentrifugation, and single-molecule fluorescence, we describe how conformational changes within NPM1 control valency and switching between the different LLPS mechanisms. We propose that this newly discovered interplay between multiple LLPS mechanisms may influence the direction of vectorial pre-ribosomal particle assembly within, and exit from the nucleolus as part of the ribosome biogenesis process.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
