

Feedback determines the structure of correlated variability in primary visual cortex
- PMID: 29483663
- PMCID: PMC5876152
- DOI: 10.1038/s41593-018-0089-1
Feedback determines the structure of correlated variability in primary visual cortex
Abstract
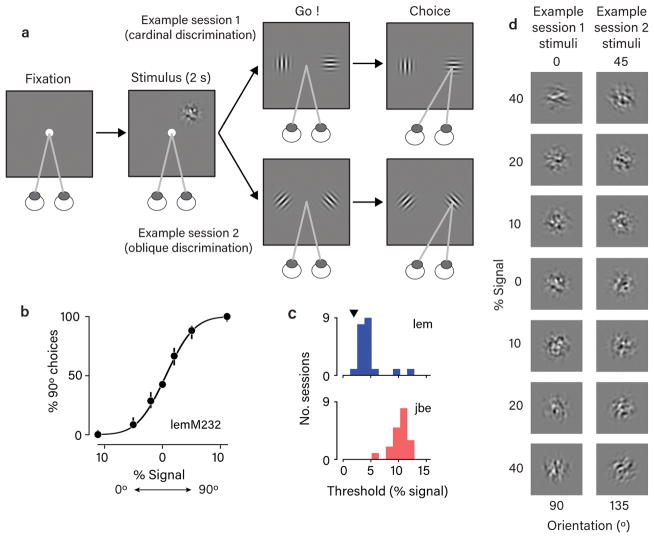
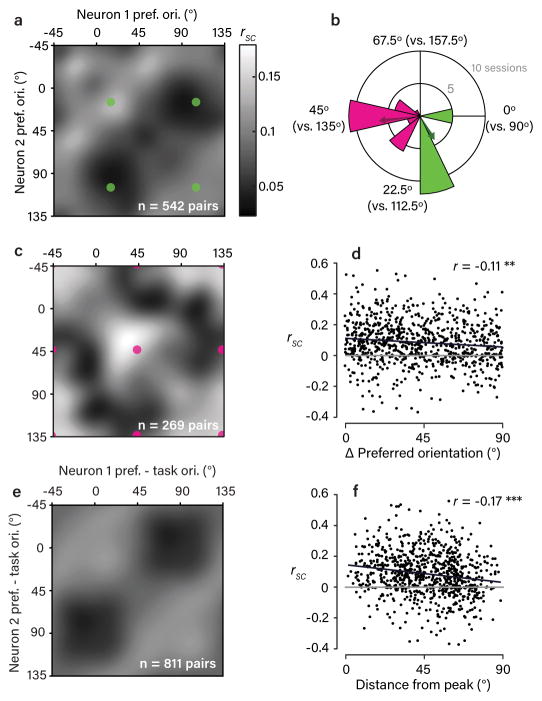
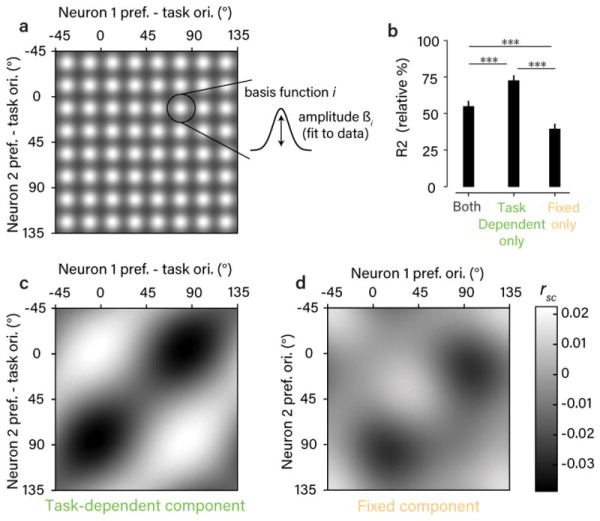
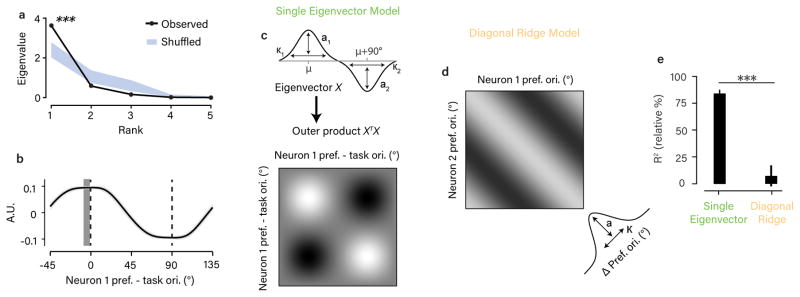
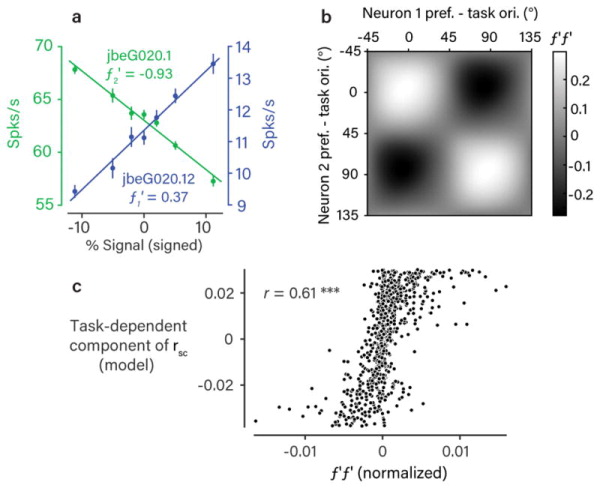
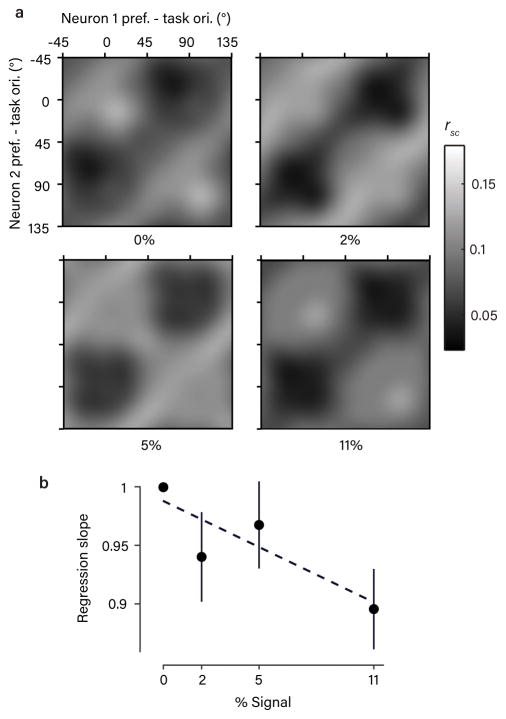
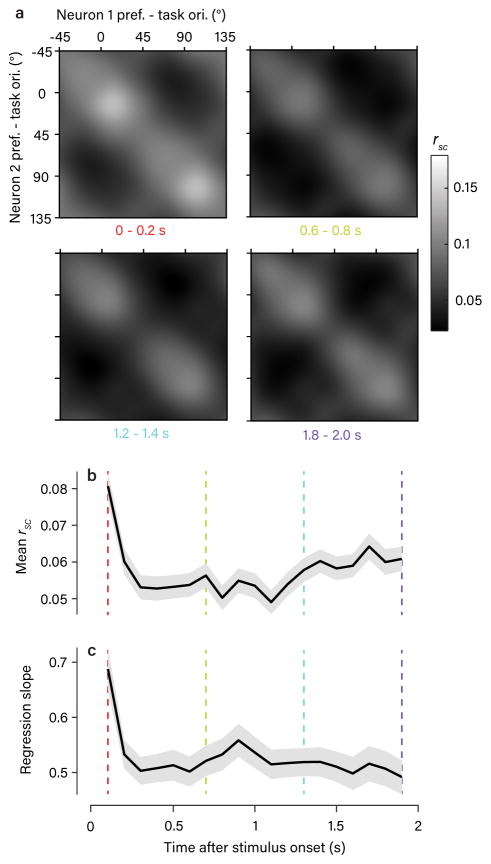
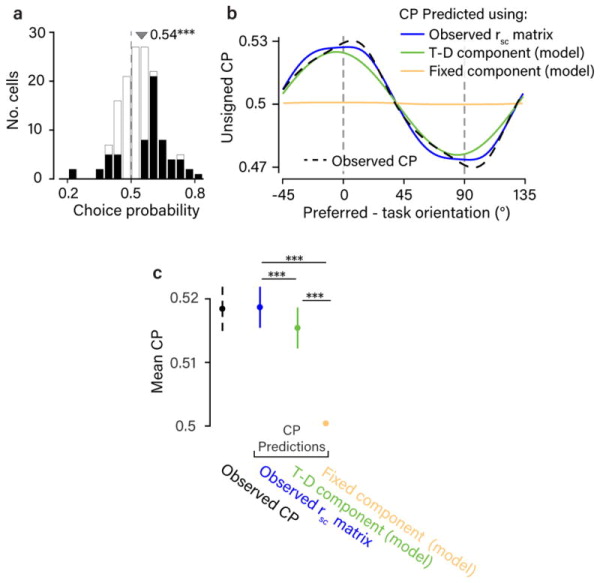
The variable responses of sensory neurons tend to be weakly correlated (spike-count correlation, rsc). This is widely thought to reflect noise in shared afferents, in which case rsc can limit the reliability of sensory coding. However, it could also be due to feedback from higher-order brain regions. Currently, the relative contributions of these sources are unknown. We addressed this by recording from populations of V1 neurons in macaques performing different discrimination tasks involving the same visual input. We found that the structure of rsc (the way rsc varied with neuronal stimulus preference) changed systematically with task instruction. Therefore, even at the earliest stage in the cortical visual hierarchy, rsc structure during task performance primarily reflects feedback dynamics. Consequently, previous proposals for how rsc constrains sensory processing need not apply. Furthermore, we show that correlations between the activity of single neurons and choice depend on feedback engaged by the task.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Tomko GJ, Crapper DR. Neuronal variability: non-stationary responses to identical visual stimuli. Brain Res. 1974;79:405–418. - PubMed
-
- Henry GH, Bishop PO, Tupper RM, Dreher B. Orientation specificity and response variability of cells in the striate cortex. Vision Res. 1973;13:1771–1779. - PubMed
-
- Sompolinsky H, Yoon H, Kang K, Shamir M. Population coding in neuronal systems with correlated noise. Phys RevE. 2001;64:51904. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
