

Parallel assembly of actin and tropomyosin, but not myosin II, during de novo actin filament formation in live mice
- PMID: 29487177
- PMCID: PMC6518266
- DOI: 10.1242/jcs.212654
Parallel assembly of actin and tropomyosin, but not myosin II, during de novo actin filament formation in live mice
Abstract
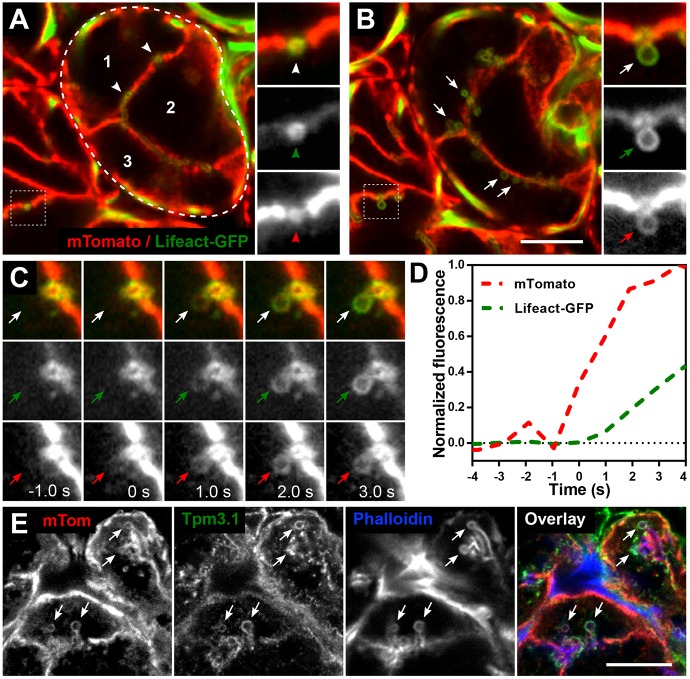
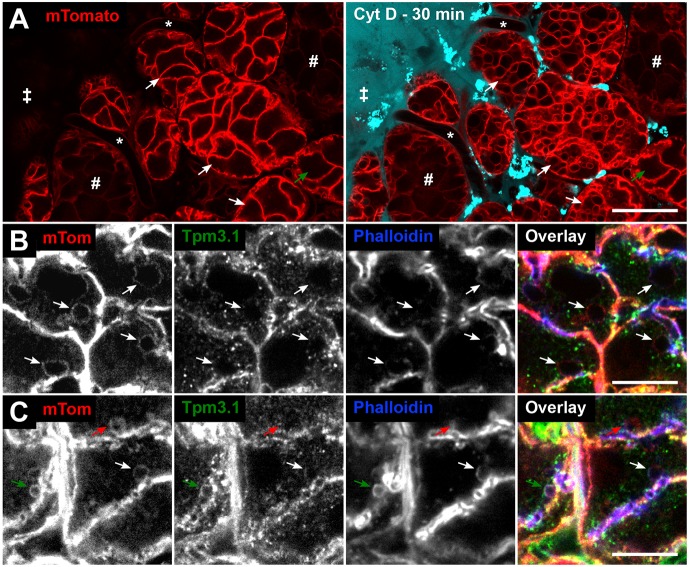
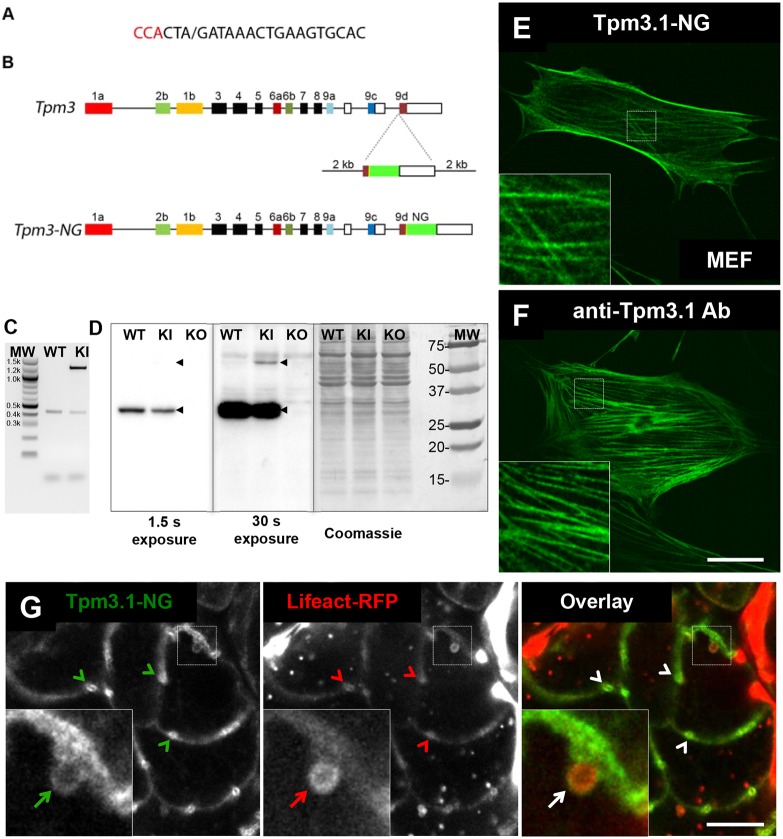
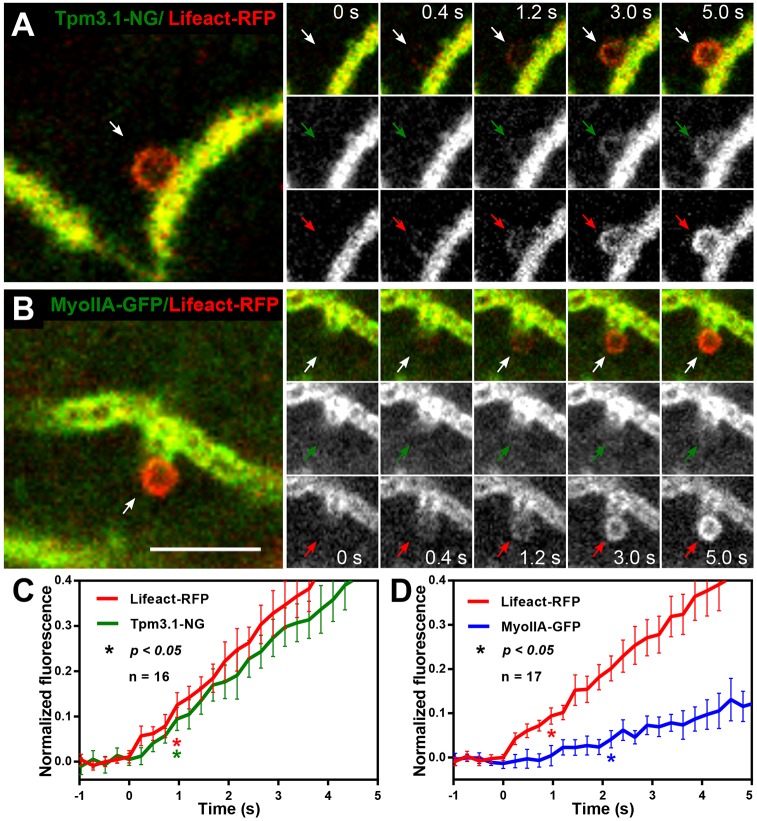
Many actin filaments in animal cells are co-polymers of actin and tropomyosin. In many cases, non-muscle myosin II associates with these co-polymers to establish a contractile network. However, the temporal relationship of these three proteins in the de novo assembly of actin filaments is not known. Intravital subcellular microscopy of secretory granule exocytosis allows the visualisation and quantification of the formation of an actin scaffold in real time, with the added advantage that it occurs in a living mammal under physiological conditions. We used this model system to investigate the de novo assembly of actin, tropomyosin Tpm3.1 (a short isoform of TPM3) and myosin IIA (the form of non-muscle myosin II with its heavy chain encoded by Myh9) on secretory granules in mouse salivary glands. Blocking actin polymerization with cytochalasin D revealed that Tpm3.1 assembly is dependent on actin assembly. We used time-lapse imaging to determine the timing of the appearance of the actin filament reporter LifeAct-RFP and of Tpm3.1-mNeonGreen on secretory granules in LifeAct-RFP transgenic, Tpm3.1-mNeonGreen and myosin IIA-GFP (GFP-tagged MYH9) knock-in mice. Our findings are consistent with the addition of tropomyosin to actin filaments shortly after the initiation of actin filament nucleation, followed by myosin IIA recruitment.
Keywords: Actin; Assembly kinetics; Cytoskeleton; Intravital; Myosin II; Tropomyosin.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Appaduray M. A., Masedunskas A., Bryce N. S., Lucas C. A., Warren S. C., Timpson P., Stear J. H., Gunning P. W. and Hardeman E. C. (2016). Recruitment kinetics of tropomyosin Tpm3.1 to actin filament bundles in the cytoskeleton is independent of actin filament kinetics. PLoS ONE 11, e0168203 10.1371/journal.pone.0168203 - DOI - PMC - PubMed
-
- Bryce N. S., Schevzov G., Ferguson V., Percival J. M., Lin J. J., Matsumura F., Bamburg J. R., Jeffrey P. L., Hardeman E. C., Gunning P. et al. (2003). Specification of actin filament function and molecular composition by tropomyosin isoforms. Mol. Biol. Cell 14, 1002-1016. 10.1091/mbc.E02-04-0244 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
