

Evidence that DNA polymerase δ contributes to initiating leading strand DNA replication in Saccharomyces cerevisiae
- PMID: 29487291
- PMCID: PMC5829166
- DOI: 10.1038/s41467-018-03270-4
Evidence that DNA polymerase δ contributes to initiating leading strand DNA replication in Saccharomyces cerevisiae
Abstract
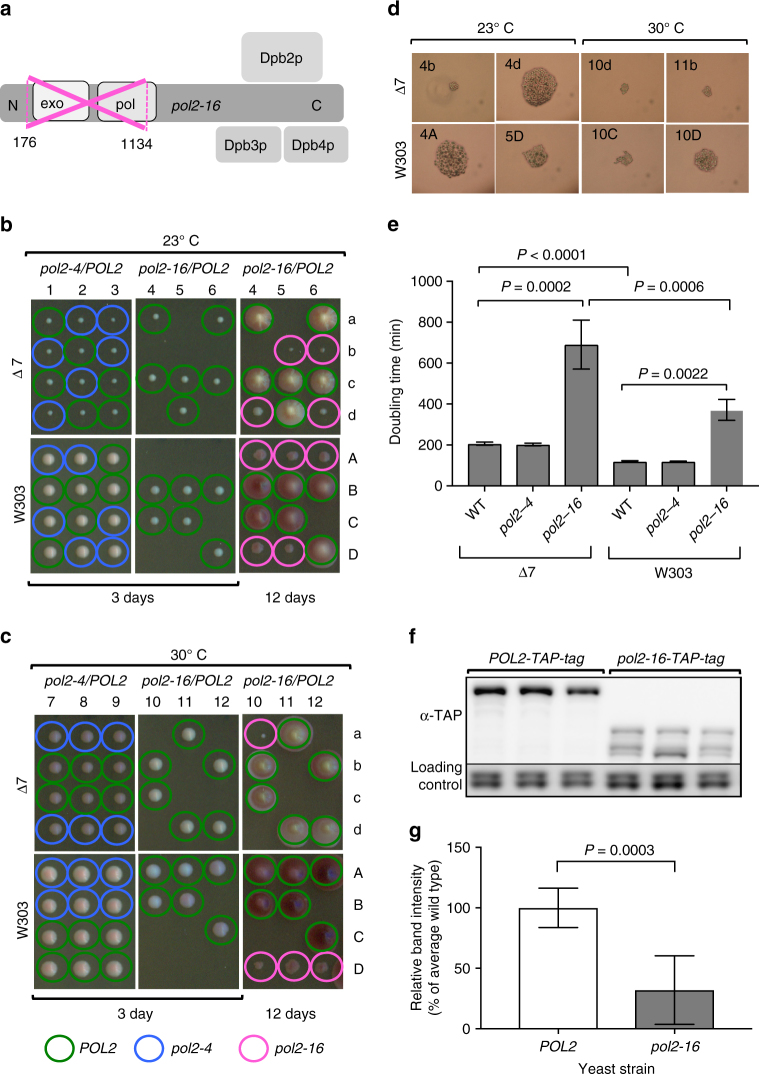
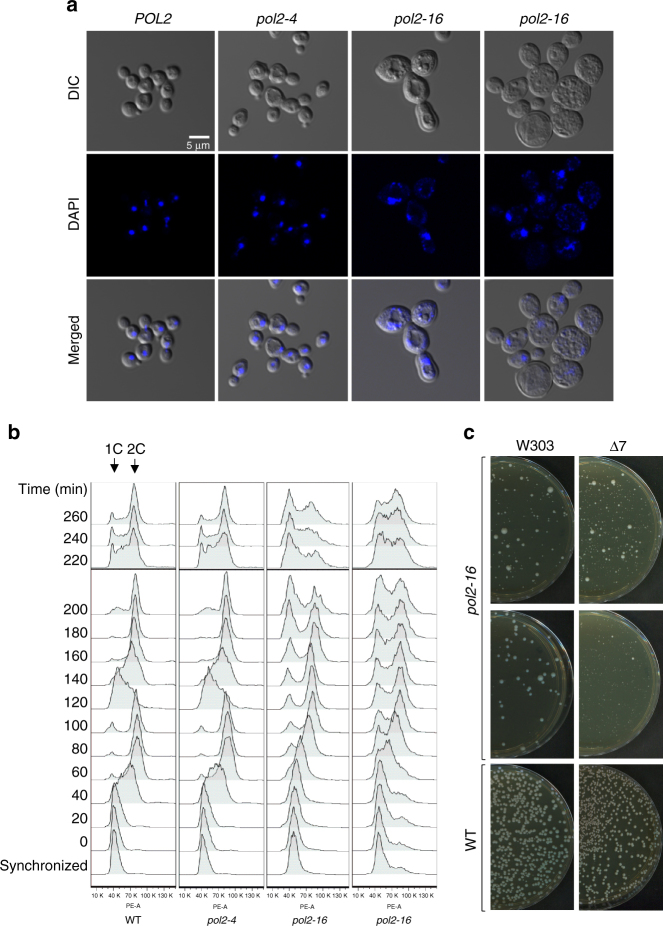
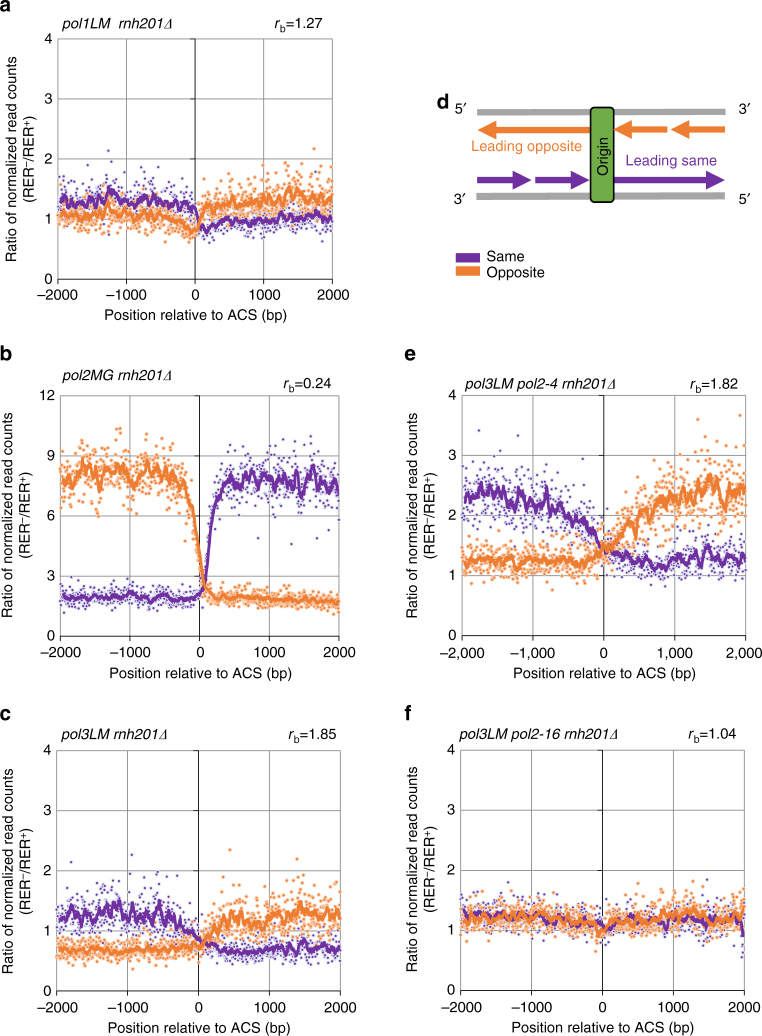
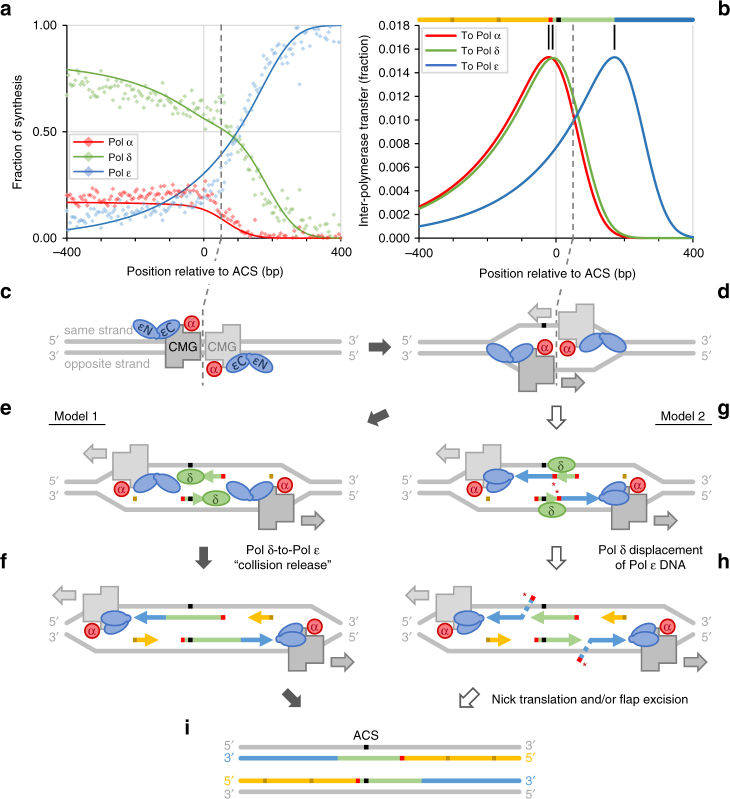
To investigate nuclear DNA replication enzymology in vivo, we have studied Saccharomyces cerevisiae strains containing a pol2-16 mutation that inactivates the catalytic activities of DNA polymerase ε (Pol ε). Although pol2-16 mutants survive, they present very tiny spore colonies, increased doubling time, larger than normal cells, aberrant nuclei, and rapid acquisition of suppressor mutations. These phenotypes reveal a severe growth defect that is distinct from that of strains that lack only Pol ε proofreading (pol2-4), consistent with the idea that Pol ε is the major leading-strand polymerase used for unstressed DNA replication. Ribonucleotides are incorporated into the pol2-16 genome in patterns consistent with leading-strand replication by Pol δ when Pol ε is absent. More importantly, ribonucleotide distributions at replication origins suggest that in strains encoding all three replicases, Pol δ contributes to initiation of leading-strand replication. We describe two possible models.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Stodola, J. L. & Burgers, P. M. Mechanism of lagging-strand DNA replication in eukaryotes. Adv. Exp. Med. Biol.1042, 117-133 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
