

Long non-coding RNA CASC15 regulates gastric cancer cell proliferation, migration and epithelial mesenchymal transition by targeting CDKN1A and ZEB1
- PMID: 29489064
- PMCID: PMC5983148
- DOI: 10.1002/1878-0261.12187
Long non-coding RNA CASC15 regulates gastric cancer cell proliferation, migration and epithelial mesenchymal transition by targeting CDKN1A and ZEB1
Abstract
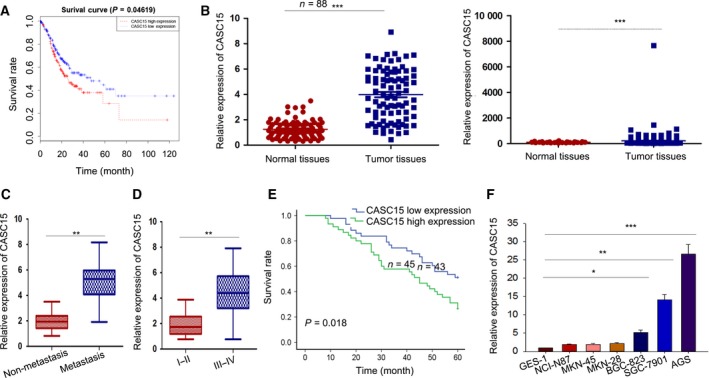
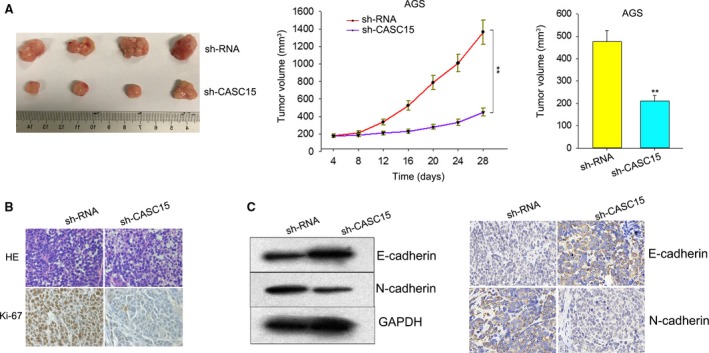
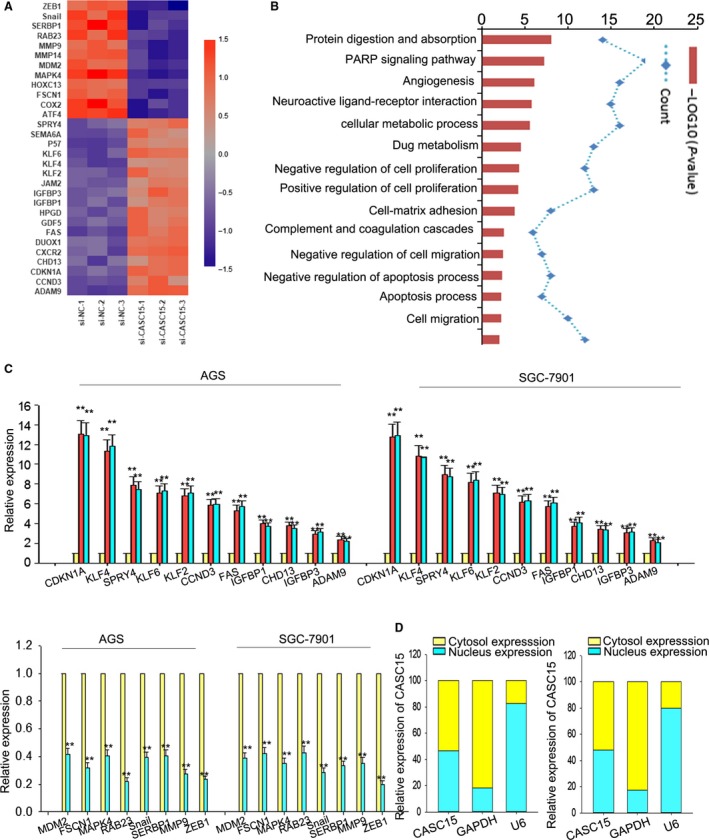
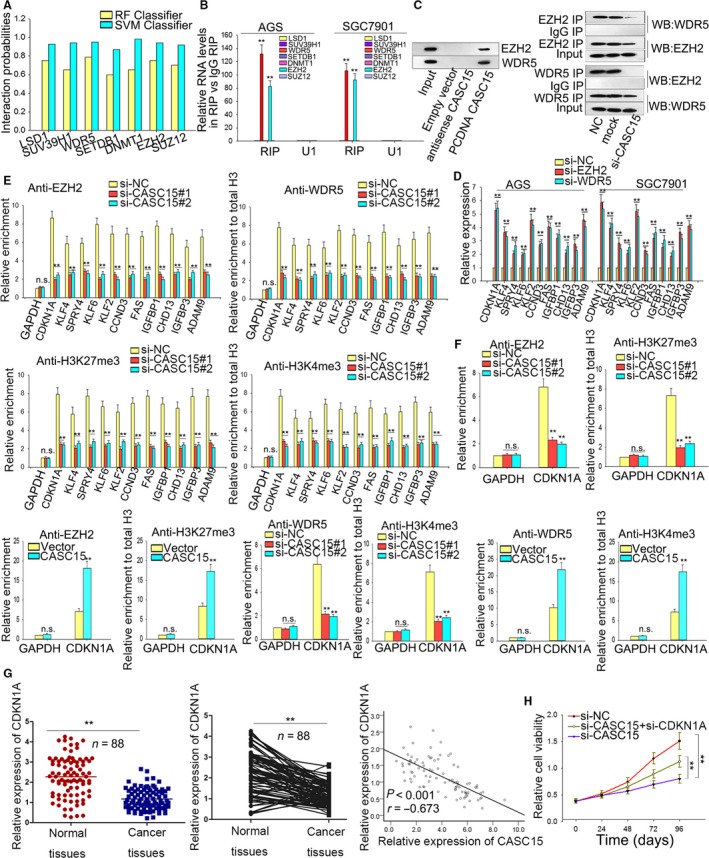
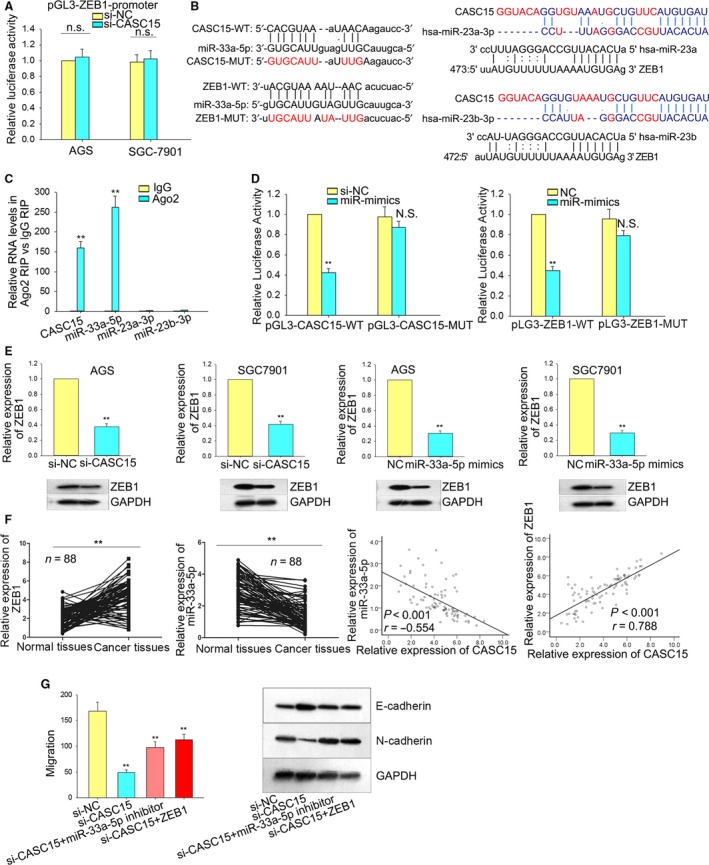
Long non-coding RNA (lncRNA) is responsible for a diverse range of cellular functions, such as transcriptional and translational regulation and variance in gene expression. The lncRNA CASC15 (cancer susceptibility candidate 15) is a long intergenic non-coding RNA (lincRNA) locus in chromosome 6p22.3. Previous research shows that lncRNA CASC15 is implicated in the biological behaviors of several cancers such as neuroblastoma and melanoma. Here, we aimed to explore in detail how CASC15 contributes to the growth of gastric cancer (GC). As predicted, the expression of CASC15 was enriched in GC tissues and cell lines as compared with healthy tissues and cells using qRT-PCR. The Kaplan-Meier method was used to demonstrate that high expression of CASC15 is linked to a poor prognosis for patients suffering from GC. Additionally, functional experiments proved that the down- or up-regulation of CASC15 inhibited or facilitated cell proliferation via the induction of cell cycle arrest and apoptosis, and also suppressed or accelerated cell migration and invasion by affecting the progression of the epithelial-to-mesenchymal transition (EMT). In vivo experiments showed that the knockdown of CASC15 lessened the tumor volume and weight and influenced the EMT process. This was confirmed by western blot assays and immunohistochemistry, indicating impaired metastatic ability in nude mice. CASC15 involvement in the tumorigenesis of GC occurs when CASC15 interacts with EZH2 and WDR5 to modulate CDKN1A in nucleus. Additionally, the knockdown of CASC15 triggered the silencing of ZEB1 in cytoplasm, which was shown to be associated with the competitive binding of CASC15 to miR-33a-5p.
Keywords: CDKN1A; ZEB1; cancer susceptibility candidate 15; epithelial-to-mesenchymal transition; gastric cancer.
© 2018 The Authors. Published by FEBS Press and John Wiley & Sons Ltd.
Figures

References
-
- Abdel‐Latif MM, Windle HJ, Davies A, Volkov Y and Kelleher D (2009) A new mechanism of gastric epithelial injury induced by acid exposure: the role of Egr‐1 and ERK signaling pathways. J Cell Biochem 108, 249–260. - PubMed
-
- Cao R, Wang L, Wang H, Xia L, Erdjument‐Bromage H, Tempst P, Jones RS and Zhang Y (2002) Role of histone H3 lysine 27 methylation in Polycomb‐group silencing. Science 298, 1039–1043. - PubMed
-
- Chaffer CL and Weinberg RA (2011) A perspective on cancer cell metastasis. Science 331, 1559–1564. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
