

Hierarchical neural architecture underlying thirst regulation
- PMID: 29489747
- PMCID: PMC6086126
- DOI: 10.1038/nature25488
Hierarchical neural architecture underlying thirst regulation
Abstract
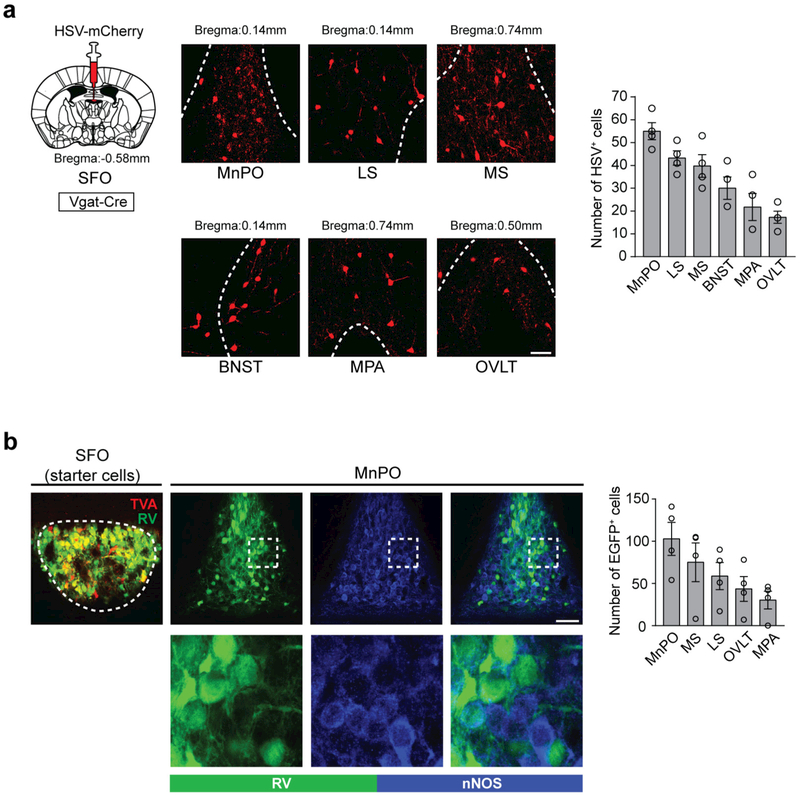
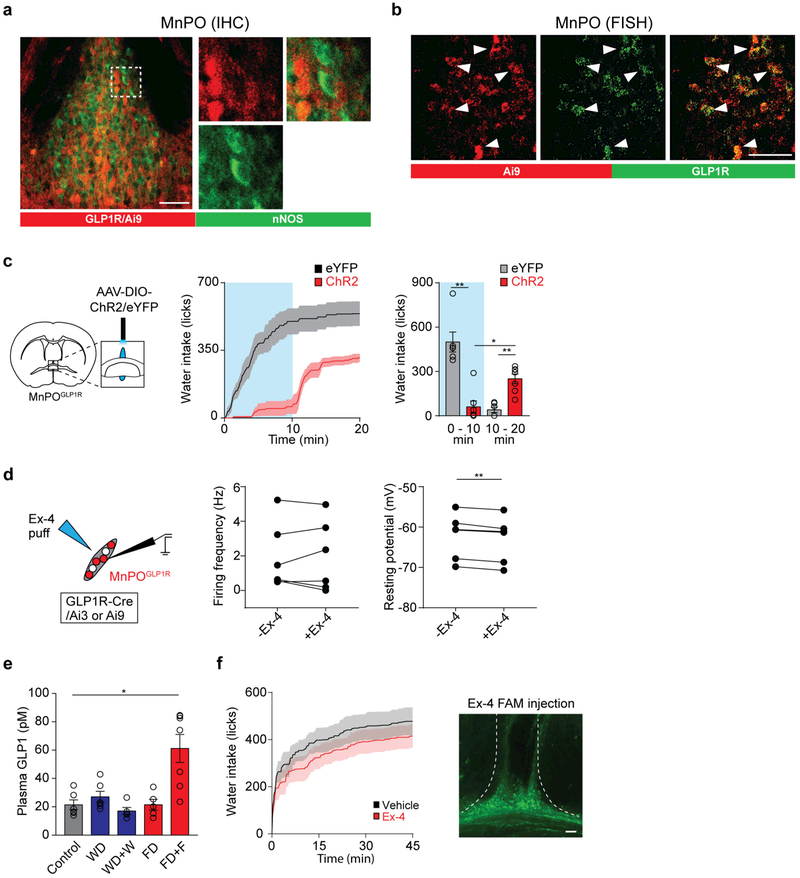
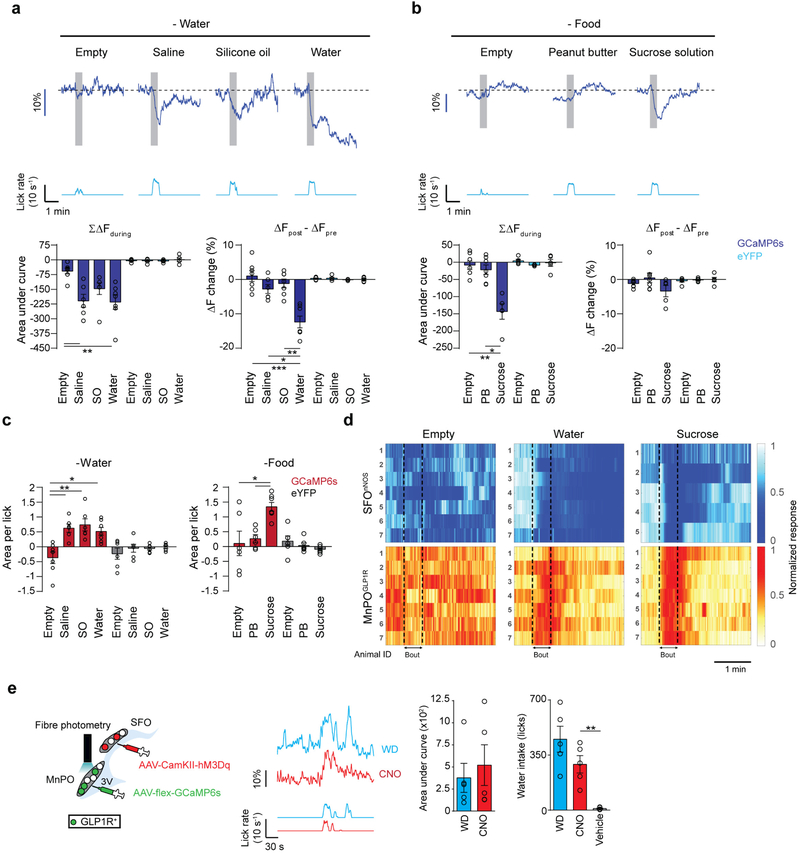
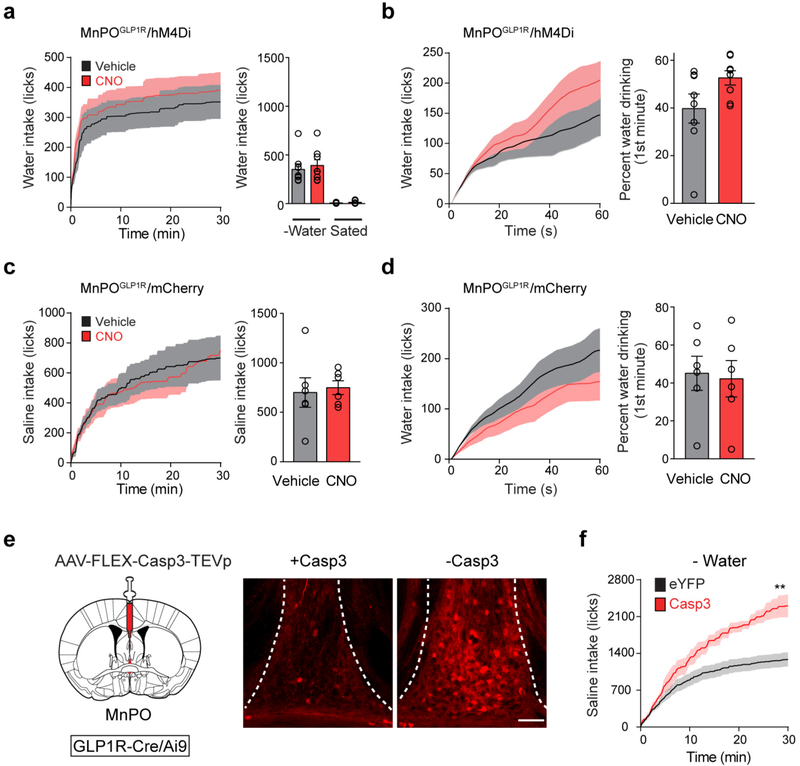
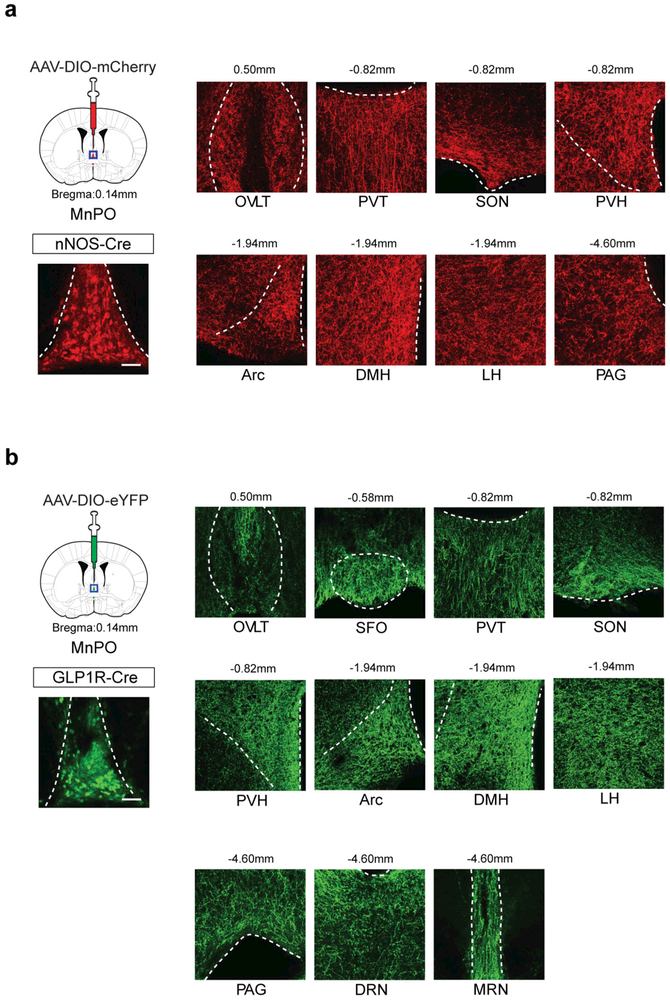
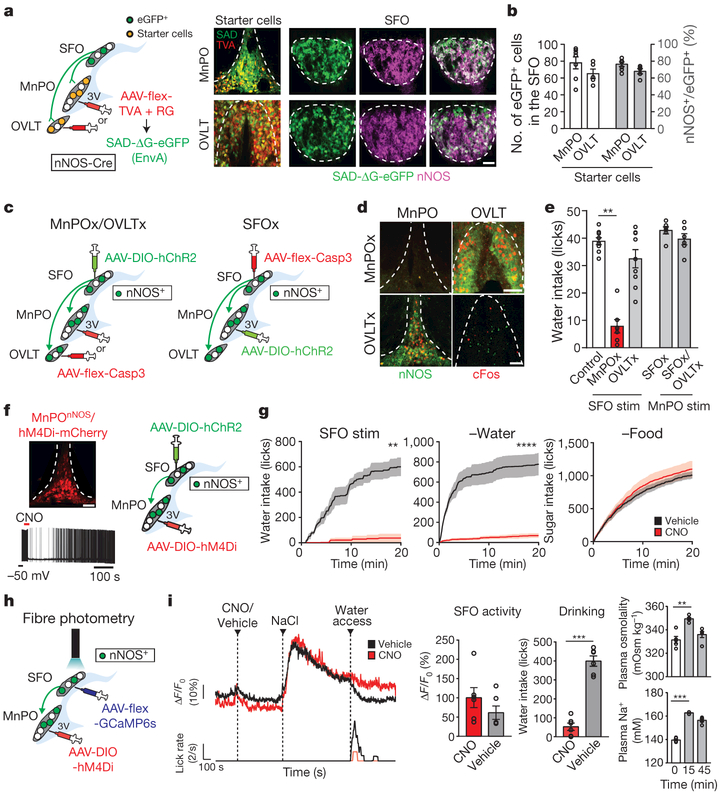
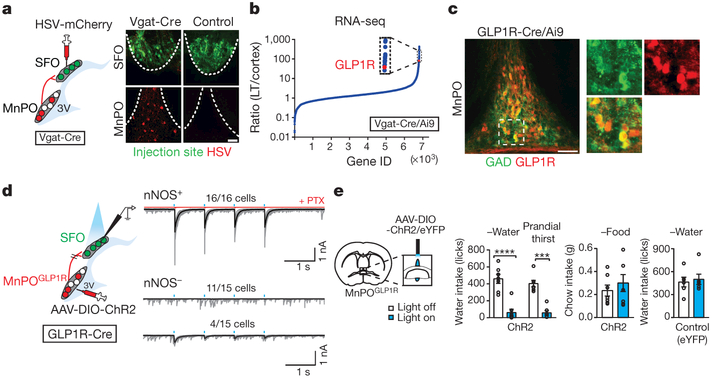
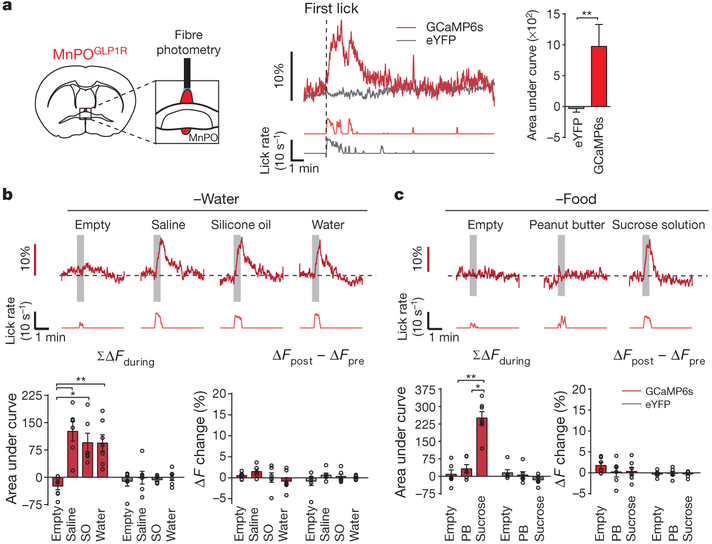
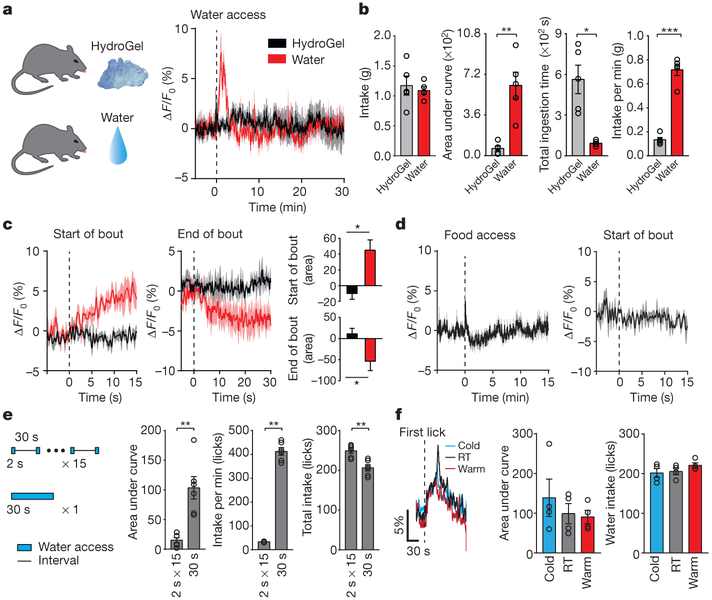
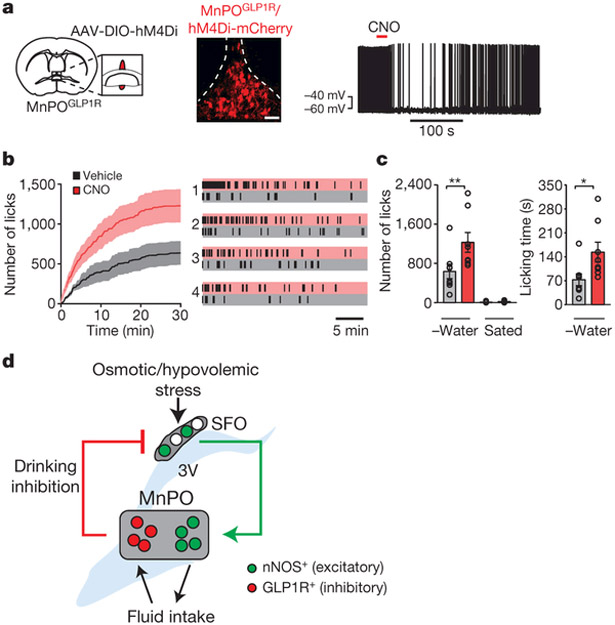
Neural circuits for appetites are regulated by both homeostatic perturbations and ingestive behaviour. However, the circuit organization that integrates these internal and external stimuli is unclear. Here we show in mice that excitatory neural populations in the lamina terminalis form a hierarchical circuit architecture to regulate thirst. Among them, nitric oxide synthase-expressing neurons in the median preoptic nucleus (MnPO) are essential for the integration of signals from the thirst-driving neurons of the subfornical organ (SFO). Conversely, a distinct inhibitory circuit, involving MnPO GABAergic neurons that express glucagon-like peptide 1 receptor (GLP1R), is activated immediately upon drinking and monosynaptically inhibits SFO thirst neurons. These responses are induced by the ingestion of fluids but not solids, and are time-locked to the onset and offset of drinking. Furthermore, loss-of-function manipulations of GLP1R-expressing MnPO neurons lead to a polydipsic, overdrinking phenotype. These neurons therefore facilitate rapid satiety of thirst by monitoring real-time fluid ingestion. Our study reveals dynamic thirst circuits that integrate the homeostatic-instinctive requirement for fluids and the consequent drinking behaviour to maintain internal water balance.
Figures
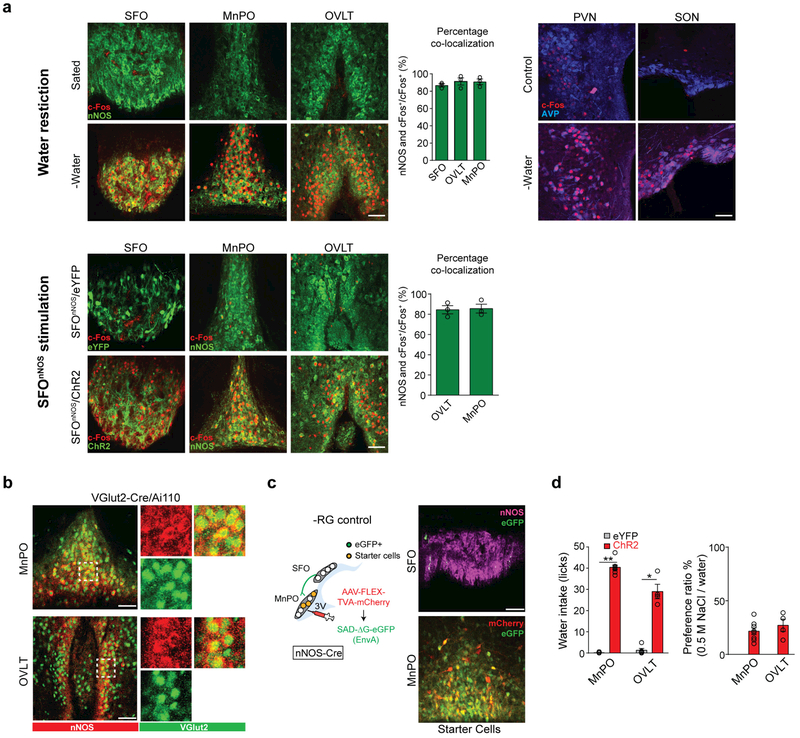
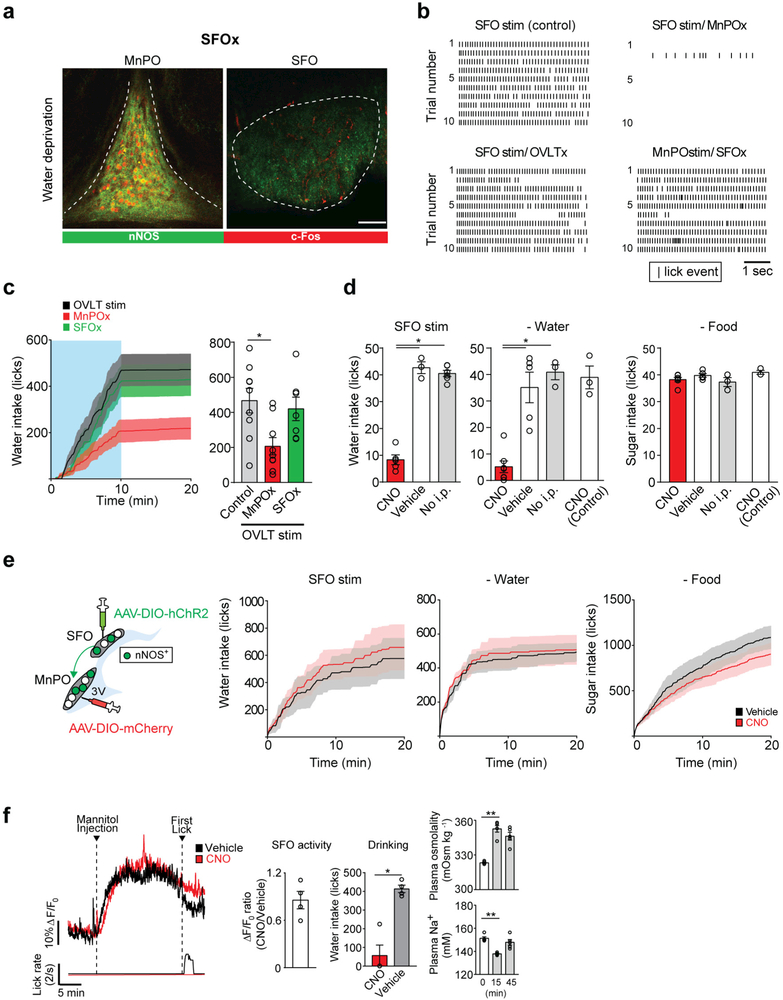
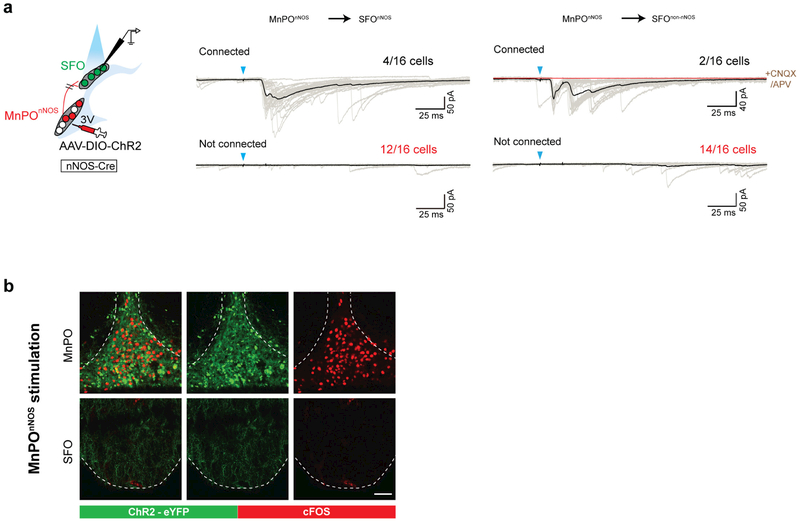
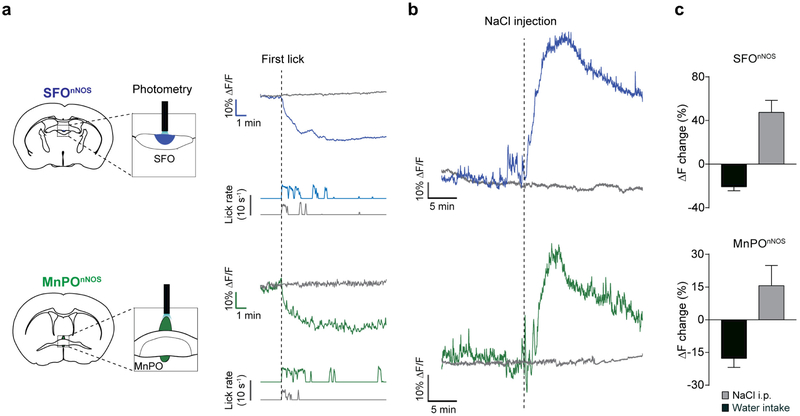
References
-
- Ramsay DJ & Booth D (eds) Thirst: Physiological and Psychological Aspects. Ch. 5, 6, 9-12, 19 (Springer, 1991).
-
- Bourque CW Central mechanisms of osmosensation and systemic osmoregulation. Nat. Rev. Neurosci 9, 519–531 (2008). - PubMed
-
- Fitzsimons JT Angiotensin, thirst, and sodium appetite. Physiol. Rev 78, 583–686 (1998). - PubMed
-
- McKinley MJ & Johnson AK The physiological regulation of thirst and fluid intake. News Physiol. Sci 19, 1–6 (2004). - PubMed
-
- Johnson AK & Gross P M. Sensory circumventricular organs and brain homeostatic pathways. FASEB J 7, 678–686 (1993). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
