

Aspirin Recapitulates Features of Caloric Restriction
- PMID: 29490275
- PMCID: PMC5848858
- DOI: 10.1016/j.celrep.2018.02.024
Aspirin Recapitulates Features of Caloric Restriction
Abstract
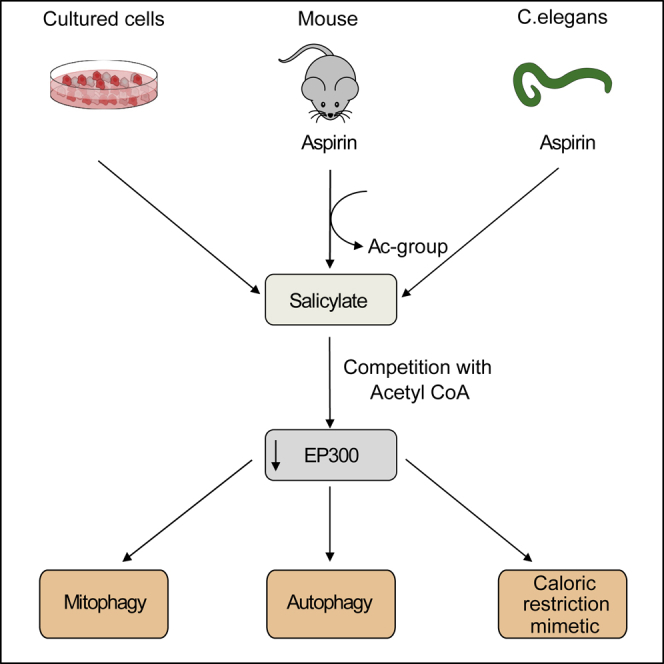
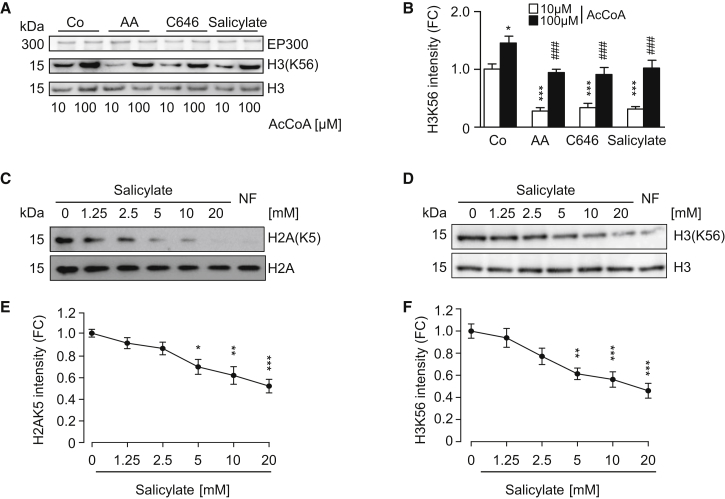
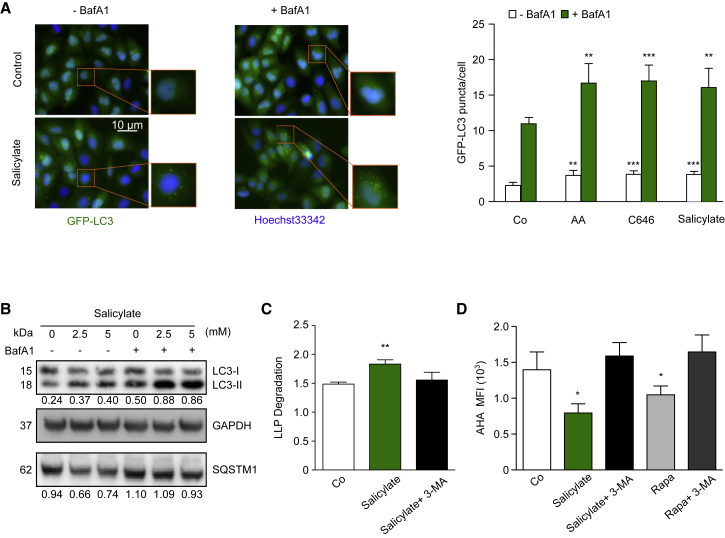
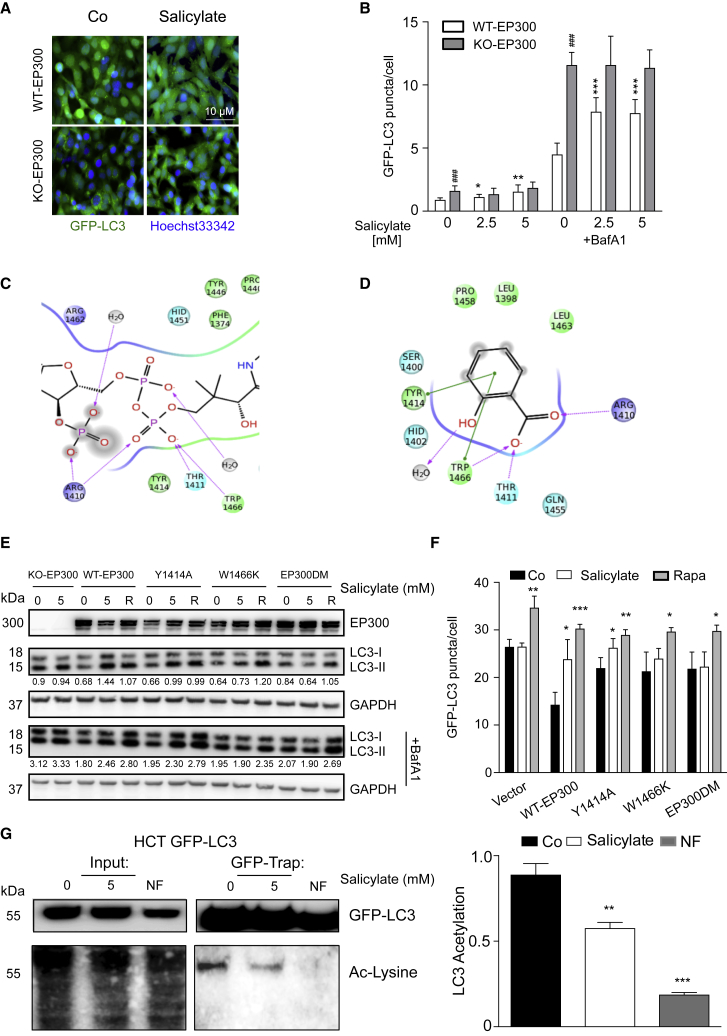
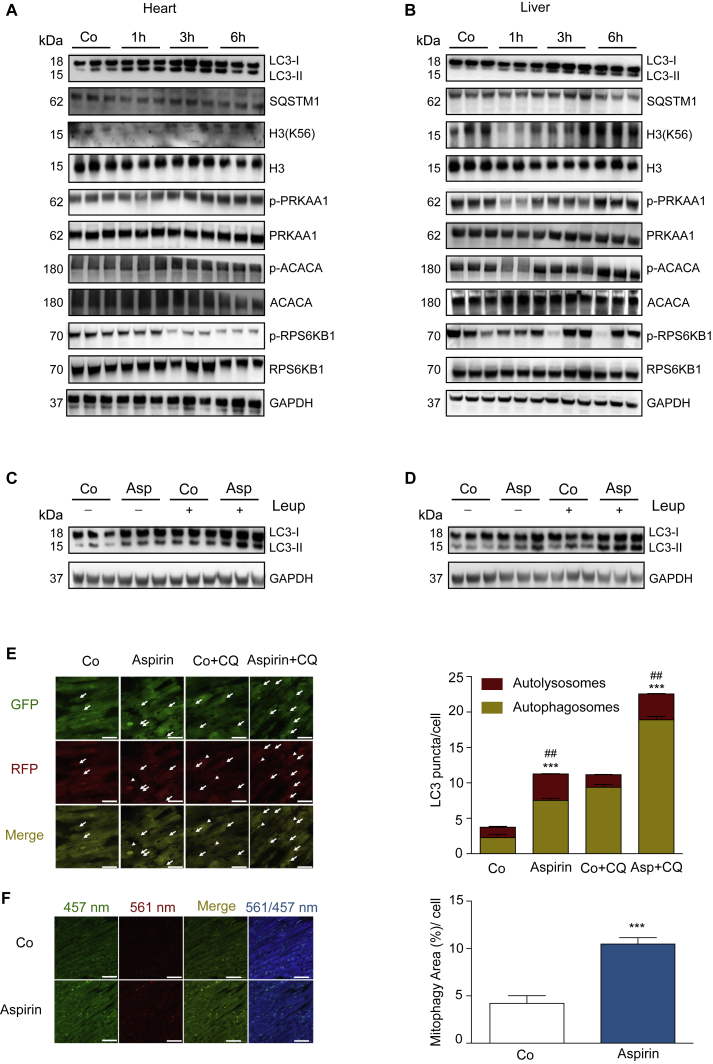
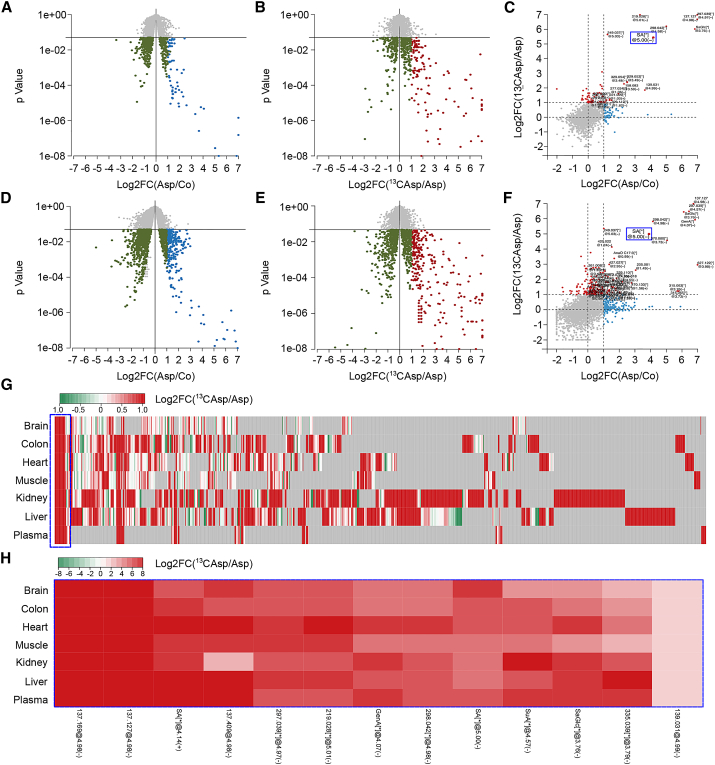
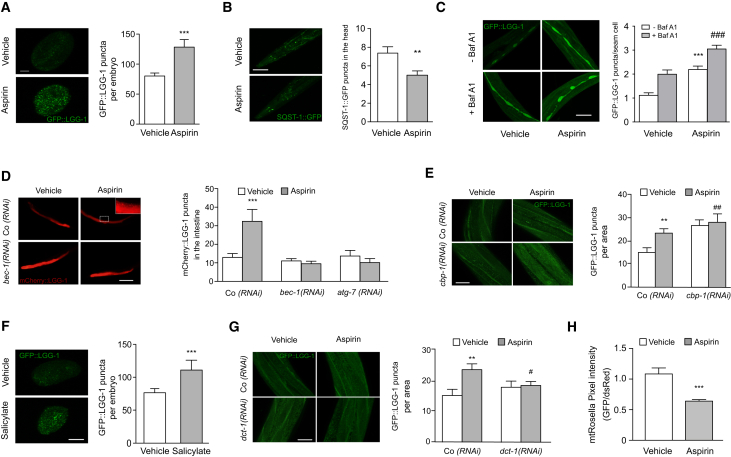
The age-associated deterioration in cellular and organismal functions associates with dysregulation of nutrient-sensing pathways and disabled autophagy. The reactivation of autophagic flux may prevent or ameliorate age-related metabolic dysfunctions. Non-toxic compounds endowed with the capacity to reduce the overall levels of protein acetylation and to induce autophagy have been categorized as caloric restriction mimetics (CRMs). Here, we show that aspirin or its active metabolite salicylate induce autophagy by virtue of their capacity to inhibit the acetyltransferase activity of EP300. While salicylate readily stimulates autophagic flux in control cells, it fails to further increase autophagy levels in EP300-deficient cells, as well as in cells in which endogenous EP300 has been replaced by salicylate-resistant EP300 mutants. Accordingly, the pro-autophagic activity of aspirin and salicylate on the nematode Caenorhabditis elegans is lost when the expression of the EP300 ortholog cpb-1 is reduced. Altogether, these findings identify aspirin as an evolutionary conserved CRM.
Keywords: EP300; acetylation; aging; autophagy; longevity; metabolome; salicylate.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Aspirin-another caloric-restriction mimetic.Autophagy. 2018;14(7):1162-1163. doi: 10.1080/15548627.2018.1454810. Epub 2018 Jul 18. Autophagy. 2018. PMID: 29929449 Free PMC article.
References
-
- Ali B., Kaur S. Mammalian tissue acetylsalicylic acid esterase(s): identification, distribution and discrimination from other esterases. J. Pharmacol. Exp. Ther. 1983;226:589–594. - PubMed
-
- Baron J.A., Cole B.F., Sandler R.S., Haile R.W., Ahnen D., Bresalier R., McKeown-Eyssen G., Summers R.W., Rothstein R., Burke C.A. A randomized trial of aspirin to prevent colorectal adenomas. N. Engl. J. Med. 2003;348:891–899. - PubMed
-
- Bauvy C., Meijer A.J., Codogno P. Assaying of autophagic protein degradation. Methods Enzymol. 2009;452:47–61. - PubMed
-
- Cantó C., Houtkooper R.H., Pirinen E., Youn D.Y., Oosterveer M.H., Cen Y., Fernandez-Marcos P.J., Yamamoto H., Andreux P.A., Cettour-Rose P. The NAD(+) precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab. 2012;15:838–847. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
