

'Cyclical Bias' in Microbiome Research Revealed by A Portable Germ-Free Housing System Using Nested Isolation
- PMID: 29491439
- PMCID: PMC5830500
- DOI: 10.1038/s41598-018-20742-1
'Cyclical Bias' in Microbiome Research Revealed by A Portable Germ-Free Housing System Using Nested Isolation
Abstract
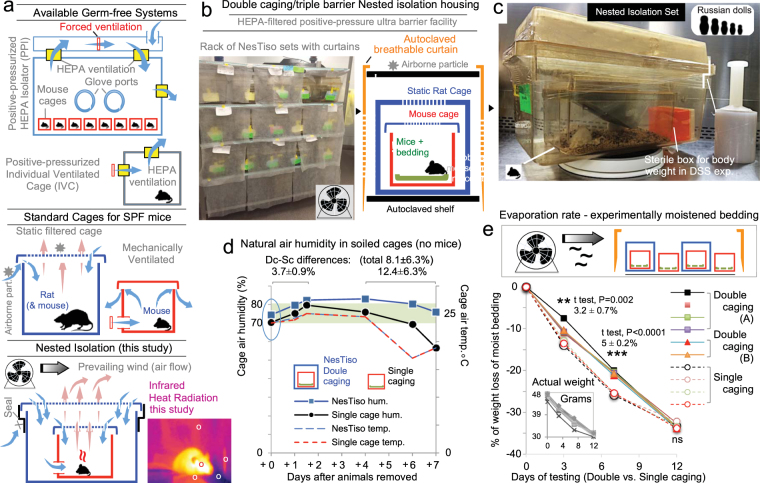
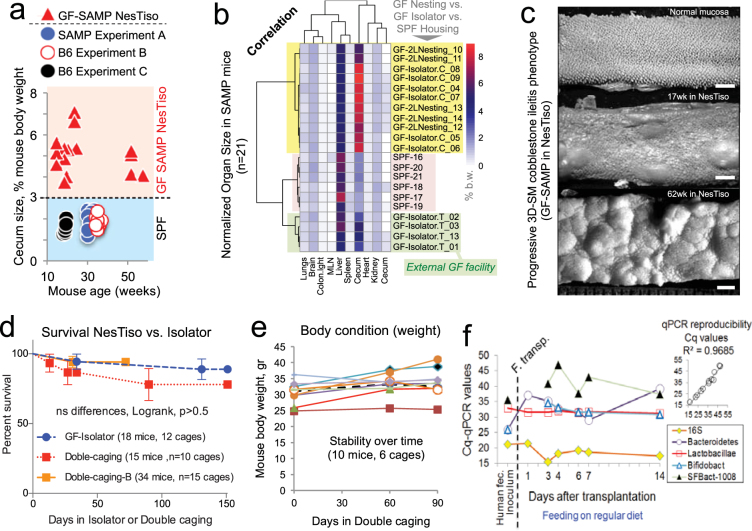
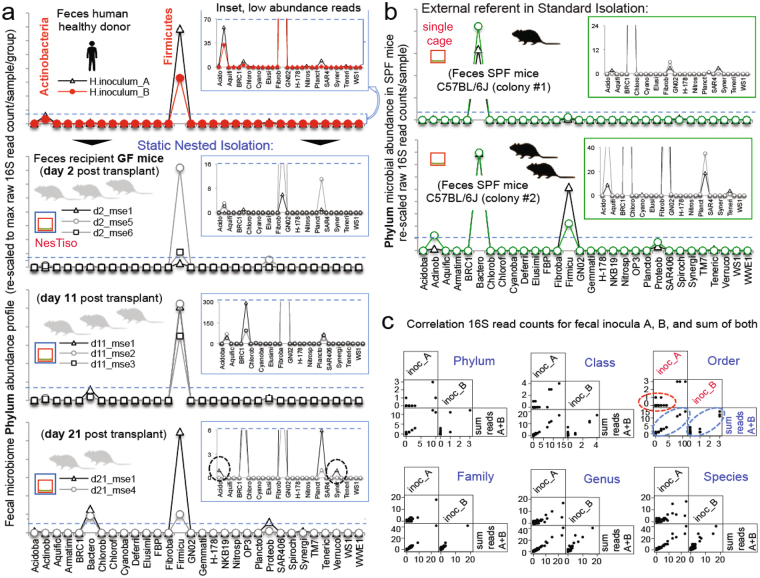
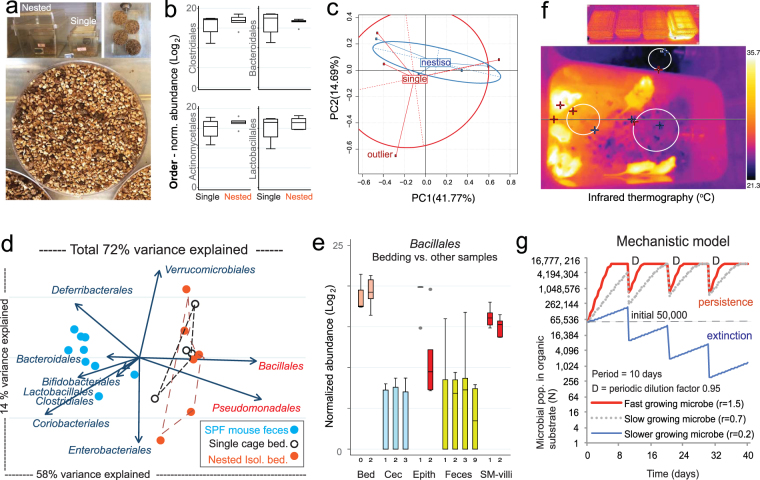
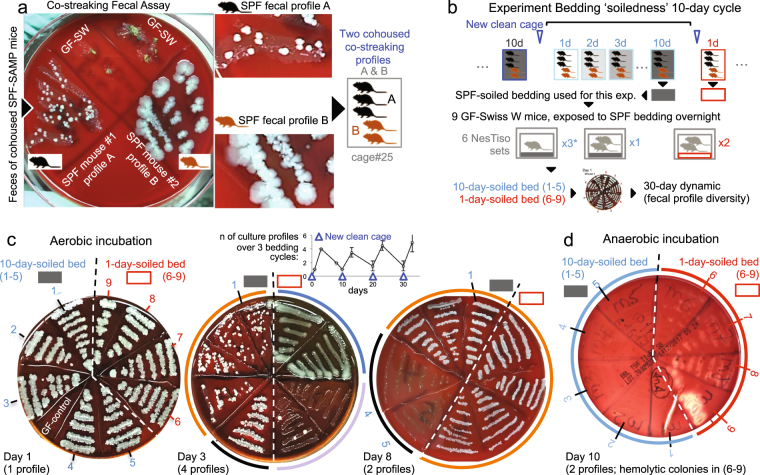
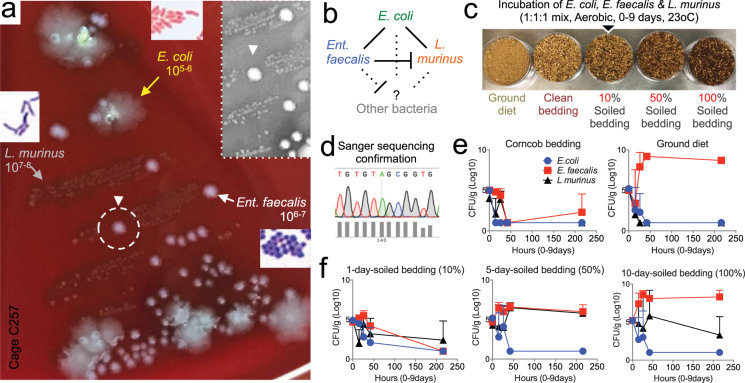
Germ-Free (GF) research has required highly technical pressurized HEPA-ventilation anchored systems for decades. Herein, we validated a GF system that can be easily implemented and portable using Nested Isolation (NesTiso). GF-standards can be achieved housing mice in non-HEPA-static cages, which only need to be nested 'one-cage-inside-another' resembling 'Russian dolls'. After 2 years of monitoring ~100,000 GF-mouse-days, NesTiso showed mice can be maintained GF for life (>1.3 years), with low animal daily-contamination-probability risk (1 every 867 days), allowing the expansion of GF research with unprecedented freedom and mobility. At the cage level, with 23,360 GF cage-days, the probability of having a cage contamination in NesTiso cages opened in biosafety hoods was statistically identical to that of opening cages inside (the 'gold standard') multi-cage pressurized GF isolators. When validating the benefits of using NesTiso in mouse microbiome research, our experiments unexpectedly revealed that the mouse fecal microbiota composition within the 'bedding material' of conventional SPF-cages suffers cyclical selection bias as moist/feces/diet/organic content ('soiledness') increases over time (e.g., favoring microbiome abundances of Bacillales, Burkholderiales, Pseudomonadales; and cultivable Enterococcus faecalis over Lactobacillus murinus and Escherichia coli), which in turn cyclically influences the gut microbiome dynamics of caged mice. Culture 'co-streaking' assays showed that cohoused mice exhibiting different fecal microbiota/hemolytic profiles in clean bedding (high-within-cage individual diversity) 'cyclically and transiently appear identical' (less diverse) as bedding soiledness increases, and recurs. Strategies are proposed to minimize this novel functional form of cyclical bedding-dependent microbiome selection bias.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
