

Accurate computational design of multipass transmembrane proteins
- PMID: 29496880
- PMCID: PMC7328376
- DOI: 10.1126/science.aaq1739
Accurate computational design of multipass transmembrane proteins
Abstract
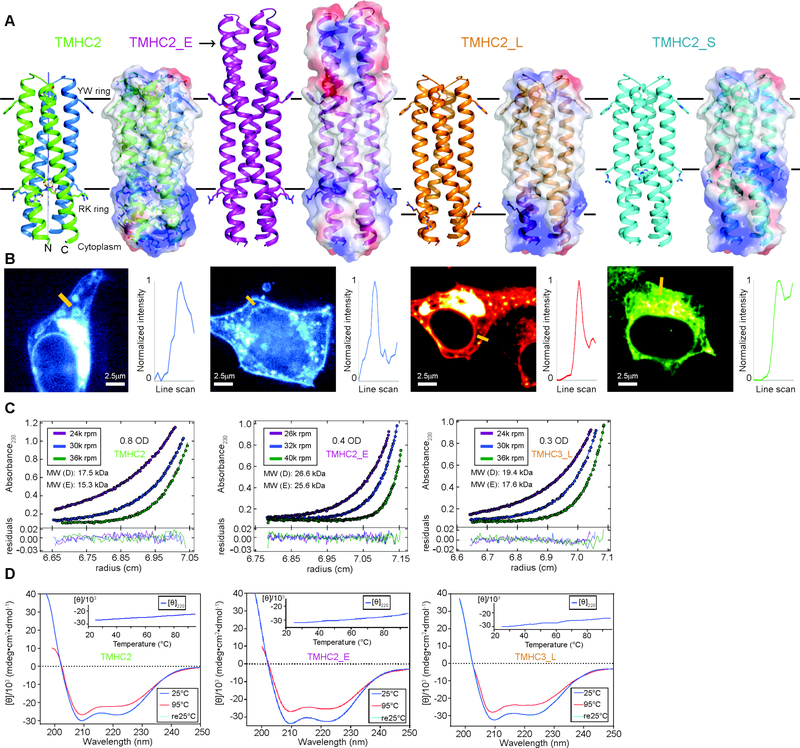
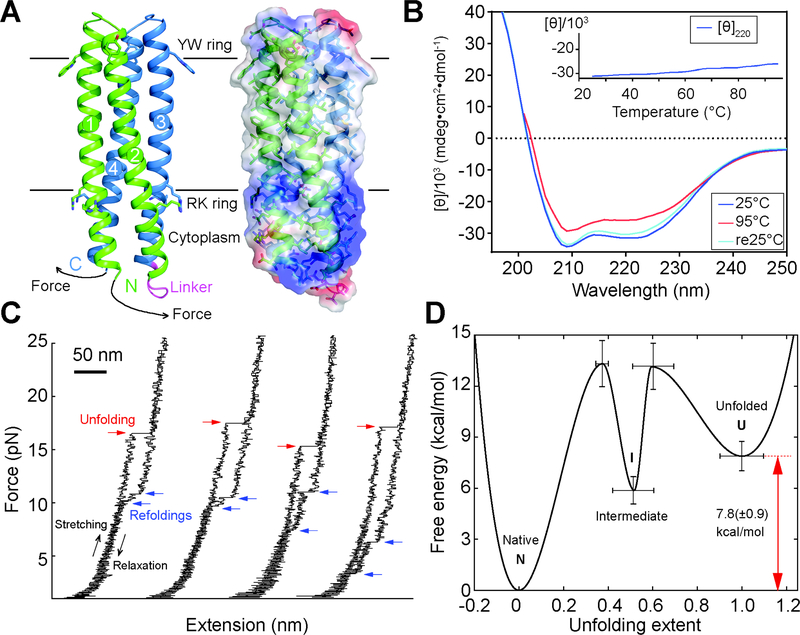
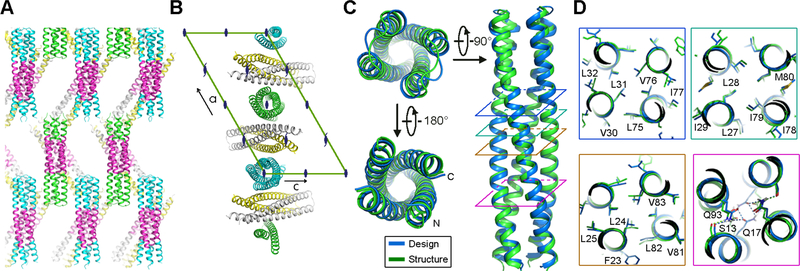
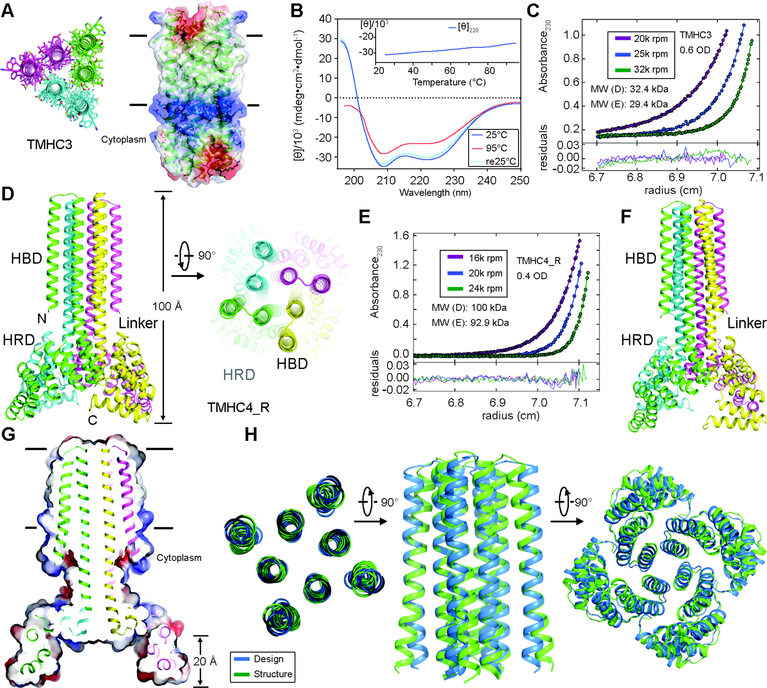
The computational design of transmembrane proteins with more than one membrane-spanning region remains a major challenge. We report the design of transmembrane monomers, homodimers, trimers, and tetramers with 76 to 215 residue subunits containing two to four membrane-spanning regions and up to 860 total residues that adopt the target oligomerization state in detergent solution. The designed proteins localize to the plasma membrane in bacteria and in mammalian cells, and magnetic tweezer unfolding experiments in the membrane indicate that they are very stable. Crystal structures of the designed dimer and tetramer-a rocket-shaped structure with a wide cytoplasmic base that funnels into eight transmembrane helices-are very close to the design models. Our results pave the way for the design of multispan membrane proteins with new functions.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Huang P-S, Boyken SE, Baker D, The coming of age of de novo protein design. Nature. 537, 320–327 (2016). - PubMed
-
- Mahendran KR et al., A monodisperse transmembrane α-helical peptide barrel. Nat. Chem. 9, 411–419 (2016). - PubMed
-
- Whitley P, Nilsson I, von Heijne G, De novo design of integral membrane proteins. Nat. Struct. Mol. Biol. 1, 858–862 (1994). - PubMed
-
- Choma C, Gratkowski H, Lear JD, DeGrado WF, Asparagine-mediated self-association of a model transmembrane helix. Nat. Struct. Biol. 7, 161–166 (2000). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
